
بافت شناسی نظری > بافت عصبی
در این صفحه می یابید:
| نورون | شکل نورون و زوائد آن | سیناپس | نوروترانسمیترها | نوروگلیا |
| اجزاء بافت شناختی مغز و نخاع | سلولهای آپاندیمی | اعصاب محیطی | عقدۀ عصبی | عصب میلینه و نامیلینه |
بافت عصبی
دستگاه عصبی تشکیل شده است از همۀ بافتهای عصبی بدن، که از لحاظ تشریحی، به دستگاه عصبی مرکزی و دستگاه عصبی محیطی تقسیم بندی میشود.
دستگاه عصبی مرکزی؛ متشکل است از مغز و نخاع که توسط جمجمه و ستون فقرات احاطه شده اند. پارانشیمِ بافت عصبی مرکزی، فاقد بافت همبند است و بر خلاف بقیۀ بافتها خالی از تارهای کلاژن و سلولهای فیبروبلاست است. البته سه پردۀ مننژ یعنی سخت شامه و نرم شامه و عنکبوتیه که مغز و نخاع را احاطه کرده اند از بافت همبند محکمی تشکیل شدهاند، همچنین عروق بزرگی که نسج مغز و نخاع را خونرسانی میکند، مانند همۀ رگهای بدن حاوی اجزاء بافت همبند هستند. نرم و شُل بودن نخاع و مغز به همین دلیل است.
شاید فایدۀ عمدهای که دستگاه عصبی مرکزی از نداشتنِ بافت پیوندی میبرد فشردگی آن باشد؛ لازمۀ انتقال پیام و پردازش آنها با سرعت بالا نزدیکی سلولهای عصبی به یکدیگر است؛ اجزاء بافت همبند، یعنی تارها و مادۀ زمینه، جای زیادی را اشغال میکند و ظاهراً ترجیح داده شده است که این فضا در اختیار نورونها و سلولهای گلیا قرار گیرد. اما اندامهای زیادی، حمایت از مغز و نخاع را از خارج، به عهده دارند؛ وجود پردههای سهگانۀ مننژ، استخوانهای محکمِ ستونِ فقرات، معماریِ خاصِ جمجمه، مایع مغزی – نخاعی، و حتی موی روی سر، از جمله عواملی هستند که به محافظت از این نسجِ نرم و آسیب پذیر کمک می کنند.
دو گروه عمدۀ سلولهای بافت عصبی عبارتاند از نورونها و سلولهای حامی یعنی سلولهای گلیا. [۱]
نوروناکثریت قریب به اتفاق نورونها قبل از تولد به وجود میآیند. نورونها بعد از بلوغ (بلوغِ نورون) قادر به تکثیر از طریق میتوز نیستند، به این ترتیب امکان ترمیم برای نورونهایی که در اثر بیماری یا ضربه یا مواد شیمیایی مثل برخی از مواد مخدر، میمیرند وجود ندارد. نورونها در حالت معمولی تا پایان عمر موجود زنده باید زنده بمانند. به این ترتیب سن بسیاری از نورونهای ما از سن شناسنامهای ما بیشتر است!
البته اضافه شدنِ تعداد نورونها پس از تولّد در انسان ادامه خواهد داشت. به نحوی که این خاصیت در انسان نسبت به بسیاری از جانوران، استثنایی و بسیار قوی است، به نحوی که در هنگام تولد، وزن مغز نوزاد، کمتر از ۴۰ درصدِ وزن مغز انسان بالغ است [۲]. وجود ملاج در سر نوزادان، امکان بزرگتر شدنِ جمجمه را به آن میدهد، و امکان بزرگ شدن جمجمه، امکان بزرگتر شدنِ مغز را فراهم میآورد [۳].
تکثیر سلولهای عصبی در انسان فقط تا چند سال پس از تولد ادامه میابد و سپس تقریباً متوقف میشود. معدودی از سلولهای بنیادین که بعد از تولد باقی میمانند، متکفل تولید نورونهای معدودی هستند که بعد از بلوغ در موجود زنده به وجود میآیند و این تعداد بسیار اندک موجب برخی از توانایی های فوقالعادۀ دستگاه عصبی میشود.
این که نورونها قدرت تکثیر ندارند، انعکاسی است از تخصصی بودنِ فوقالعادۀ آنها. نورونها برای آن که بتوانند وظیفۀ اصلی خود را انجام دهند، از بسیاری از اعمال حیاتی عاجز شده اند. برخی از اعمال حیاتی نورونها بر عهدۀ سلولهای گلیا گذاشته شده است. نورون تنها سلول متخصص نیست که برخی از تواناییهای حیاتی خود را از دست داده اند. گلبول قرمز یک مثال دیگر است که حتی هستۀ خود را فدای وظیفۀ خود کرده است.
نورونها سلولهای عموماً بزرگی هستند. اما بیشتر حجمِ آنها مربوط به استطالههای متعدد و بسیار طولانی آنهاست. یک نورونِ حسی که از انگشت پای انسان به نخاع میرود میتواند بیش از یک متر طول داشته باشد. نورونِ حسی سمِ دستِ یک زرّافه، طبعاً از این هم طولانیتر است.
|
نورون چقدر بزرگ است؟ [اگر قطر یک آکسون را ۱۰ میکرومتر فرض کنیم، و طول آن را یک متر، حجم آن چیزی حدود ۷۸۵۰۰۰۰۰ میکرومتر مکعب خواهد بود. اگر این سلول کروی شکل بود قطر آن ۵۳۱ میکرون میشد که بیش ازنیم میلیمتر است. چنین سلولی بدون میکروسکوپ و با چشم به آسانی قابل مشاهده میبود. |
نورون در هر جایی از بدن باشد برای انتقال یک “پیام” از جایی به جایی دیگرتعبیه شده است. این تعبیر ساده از کار نورون، مستلزم سه عملیاتِ مجزا از هم است؛ ۱) اخذ پیام از یک محرک خارجی یا از یک نورونِ دیگر ۲) حرکت پیام در طول نورون و استطالههای آن و ۳) تحویل دادن پیام به نورونِ دیگر یا به یک سلولِ عملگر مانند سلول عضله.
برای فهم مراحل فوق حداقل آنچه باید بدانیم عبارت است از ۱) کیفیت عکسالعمل نورن در مقابل پیامهای شیمیایی که از دیگر نورونها به آن میرسد ۲) شکل نورون و استطالههای آن و ۳) مواد شیمیایی که نورون برای ارتباط با دیگر نورونها تولید میکند که به نوروترانسمیتر موسوم است.
ما مسیر خود را در این راه از شکل نورون آغاز میکنیم:
نورون تشکیل شده است از یک جسم سلولی و استطالههای طولانی اطراف آن. این استطاله ها از لحاظ شکل و عملکرد، به دو گروه تقسیم میشوند؛ دندریت و آکسون.
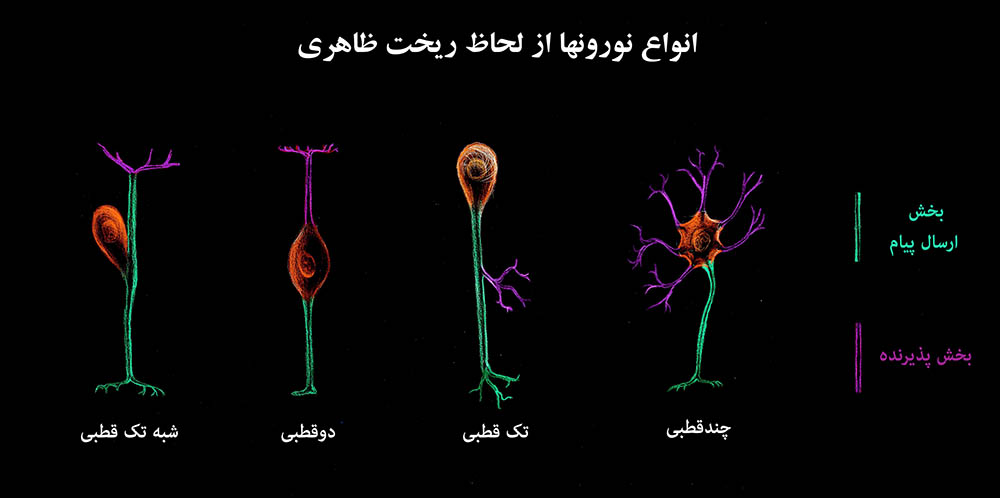
شکل ۱- ریخت شناسی نورون؛ نورون تشکیل شده است از جسم سلولی و استطالههای آن. زوائدی که مانند شاخۀ درخت از آنها جدا و منشعب میشوند و پذیرنده و آورندۀ پیام به سوی جسم سلولی هستند را دندریت نامیده اند. تار منفردی که پیام از طریق آن به سوی سلولِ دیگر میرود را آکسون نامیده اند. این قانون کلی استثنائاتی هم دارد.
نورونِ تک قطبی از اختصاصاتِ بیمهرگان است اما در دستگاه خودکار (اتونوم) پستانداران هم وجود دارد. نورون شبه تک قطبی شاخصۀ گانگلیون ریشۀ خلفی نخاع و برخی از اعصاب جمجمه ای است. مشهورترین نورونهای دو قطبی در پستانداران نورونهای موجود در شبکیۀ چشم هستند. این طرح شماتیک را با عکسهای موجود در شکلهای ۴(عکس ۴-۱)، ۱۲-الف، ۱۳، و ۱۵ مقایسه کنید.
دندریتها به همراه بخشی از خودِ جسم سلولیِ نورن، بخشِ آورانِ نورون را تشکیل میدهند. نورونها عموماً یک یا بیش از یک دندریت اولیه دارند که از جسم سلولی جدا میشود. درمقابل؛ قانون عمومی در مورد آکسون منفرد بودن آن است. هر دندریت اولیه ممکن است به دندریتهای ثانویه و ثالثیه و بیشتر منشعب شود. دندریتها ممکن است صاف یا زبر باشند و برامدگیهای میخ مانندی داشته باشند که آن ها را خار مینامیم. [۴]
تعداد سلولهای مغز را حدود ۱۰۰ میلیارد تخمین زده اند [۵] که این تعداد را میتوان حداقل به ۱۰۰۰ نوع تقسیم بندی کرد. [۶]
پس از آن که کامیللو گلژیِ ایتالیایی شیوۀ معجزه گونۀ خود را برای رنگامیزی سلولهای عصبی ابداع کرد و سانتیاگو رامون کاخالِ اسپانیایی کنجکاوی بی حد و تلاش بی وقفۀ خود را وقف مشاهدۀ دستگاه عصبی کرد، راه طبقه بندیِ نورونها بر اساس شکل ظاهری آنها باز شد. اما در دو دهۀ گذشته پیشرفتهای حوزۀ زیستشناسیِ ملکولی، تقسیم بندیِ نورونها را بر اساس کارکرد آنها بر اساس بیان ژن در آنها میسر کرده و اعتبارِ ریخت شناسی میکروسکوپی نورونها تا حدی کاهش یافته است. با این وجود هنوز هم برخی از نامگذاریهای پیشتازان عصبشناسی، معتبر باقی مانده است. دلیل اصلی بقاء این اعتبار، هماهنگی نسبیِ شکل و عملکردِ نورونها است.

به طور عمومی نورونها را از لحاظ ظاهری به نورنهای تک قطبی، دو قطبی، چند قطبی و شبه تکقطبی تقسیم بندی می کنند. نورونهای تکقطبی مشخصّۀ دستگاه عصبیِ بیمهرگان هستند اما در مهره داران، در دستگاه خودکار (اتونوم) که تنظیم اعمال حیاتیِ ابتدایی را بر عهده دارند نیز دیده میشوند. نورونهای دوقطبی، دارای یک دندریت در یک سمت و یک آکسون در سمت مقابل هستند، در شبکیۀ چشم یک طبقه از سلولهای دو قطبی وجود دارد. در دستگاه عصبی اکثریت، با نورونهای چند قطبی است. سلول پورکنژ در مخچه، (شکل ۱۳) یک سلول چند قطبی است با تعداد بسیار زیادی از نورونها که به کرّات منشعب میشوند. این انشعابهای پرشمار، امکان اتصال و سیناپس با تعداد زیادی نورون را در آن فراهم می کنند. تعداد سیناپسهای یک سلول منفردِ پورکنژ را تا یک میلیون سیناپس تخمین زده اند [۶] (این تعداد را با تعداد سلولهایی که با یک سلول، سیناپس دارند اشتباه نکنید، از هر سلولِ غیر، ممکن است صدها سیناپس با یک سلولِ پورکنژ برقرار شود.). جسمِ سلولیِ نورونهای شبه تک قطبی، در گانگلیون خلفی قرار دارند. این سلولها در دورۀ جنینی دو قطبی بوده اند و سپس در طی رشد، ریشۀ دو استطاله ای که از جسم سلولی خارج میشوند، به هم جوش خورده اند بنابراین در ابتدا دو قطبی بوده اند علیهذا به آنها شبه تک قطبی یا تک قطبیِ کاذب گفته میشود.
نورونها را میتوان از لحاظ کارکرد نیز به سه دستۀ کلی تقسیم کرد؛ نورونِ آوران، نورنِ وابران و نورونِ واسطه.
تجمع این نورونهای متنوع با کیفیتهای متنوع در جایجای مغر و نخاع، مناطقی را در دستگاه عصبی مرکزی به وجود آورده که هر یک از این مناطق یکی ازوظایف بسیارِ مغز را بر عهده دارد.
همانگونه که اشاره کردیم، از جسم سلولی، بیش از یک آکسون منشعب نمیشود، هرچند آکسون در طول مسیر خود و در انتهای خود، ممکن است شاخه هایی داشته باشد. جسم سلولی در نقطه ای که آکسون از آن خارج میشود برجسته میشود. به این برجستگی پشتۀ آکسون گفته اند. آکسون بخش وابرانِ نورون را تشکیل میدهد. برجستگیهای کوچکی در انتها یا/و در طول مسیر آکسون در سطح آن وجود دارد که به آنها، به ترتیب تکمۀ انتهایی و تکمۀ سرراهی میگوییم.
سیناپس ساختار کاملاً تخصص یافته ای است که بین تکمۀ یک نورون (نورونِ پیشسیناپسی) و نورونِ دیگر(نورونِ پسسیناپسی)، اتصال برقرار میکند. در داخل فضای آکسونِ نورونِ پیشسیناپسی و در نزدیکی سیناپس، نافه (وزیکول) های سیناسی متعدد وجود دارد. این نافه ها که دیوارۀ آنها مانند غشاء سلول از یک دولایۀ لیپیدی تشکیل شده، حاوی نوروترانسمیتر است. این نافه ها در هنگام انتقال پیام، محتوای خود که همان نوروترانسمیترها هستند را طی پدیدۀ اگزوسیتوز به فضای سیناپسی تخلیه می کنند. وصول این ملکولها به گیرنده های موجود در سطح نورونِ پسسیناپسی موجب شروع یک سلسله واکنش در غشاء نورون پسسیناپسی میشود که در نهایت به ایجاد پیام عصبی در آن میگردد.
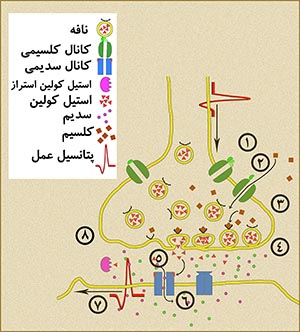 شکل ۲- سیناپس شیمیایی؛ وصولِ پتانسیل عمل به انتهای آکسون موجب تخلیۀ نوروترانسمیتر از نافۀ (وزیکول) سیناپسی به درزِ سیناپس و از آن طریق به گیرنده های سطح نورون یا سلولِ عملگرِ بعدی میشود. اتصالِ نوروترانسمیتر به گیرنده موجب ایجاد پتانسیل عمل در این سلول میشود. برای توضیح بیشتر روی عکس کلیک کنید.
شکل ۲- سیناپس شیمیایی؛ وصولِ پتانسیل عمل به انتهای آکسون موجب تخلیۀ نوروترانسمیتر از نافۀ (وزیکول) سیناپسی به درزِ سیناپس و از آن طریق به گیرنده های سطح نورون یا سلولِ عملگرِ بعدی میشود. اتصالِ نوروترانسمیتر به گیرنده موجب ایجاد پتانسیل عمل در این سلول میشود. برای توضیح بیشتر روی عکس کلیک کنید.
مواد شیمیایی رها شده در درز سیناپسی به سرعت توسط آنزیمهای موجود در این فضا شکسته میشوند یا/و مجدداً توسط نورونِ پیشسیناپسی بازجذب میشود و در درونِ نافه ها برای استفاده در انتقال پالسِ بعدی ذخیره میشود. همانطور که میبینیم، غشاء نورونهای پیش- و پسسیناپسی، در ناحیۀ سیناپس وظایف تخصصی پیچیده ای دارند. این پیچیدگی در ساختار غشاءهای این دو قسمت به صورت تغییراتی که در تراکم غشاء سلولی در ناحیۀ سیناپس دیده میشود، انعکاس یافته است. این اختلاف تراکم در غشاء فقط با میکروسکوپ الکترونی قابل مشاهده است. در سیناپسهای تحریکی، ضخامت این دو تراکم به یک اندازه است و از این جهت سیناپس، متقارن است. در سیناپسهای کاهنده، ضخامت تراکم پس سیناپسی از آنِ پیش سیناپسی، بیشتر است و سیناپس از این نظر نامتقارن است.[۷]
تعداد قلیلی از سیناپسها بدون استفاده از مواد شیمیایی پیام خود را به سلولِ پس-سیناپسی، از طریق پیام الکتریکی منتقل می کنند.
هنگامی که پیام عصبی به هر طریقی، به یک نورون برسد، به شکل یک پیام الکتریکی، طول نورون را از ابتدا تا انتهای مسیر خود را طی میکند. آنچه در طول یک نورون و زوائد آن منتقل میشود، یک موج الکتریکی است. اما هنگامی که به انتهای مسیر خود در آکسون میرسد، به صورت شیمیایی ترجمه میشود و سپس به عصب بعدی انتقال مییابد، که در بالا به اختصار به آن اشاره شد. هر نورون تنها از یک نوروترانسمیتر برای انتقال پیام خود به سلول بعد از خود استفاده میکند. نوروترانسمیترهای مهم مورد استفاده در بدن انسان عبارتاند از اِل-گلوتامات، گابا (گاما آمینو بوتیریک اسید)، دوپامین، سروتونین، استیلکولین، نورادرنالین و گلیسین. به همراه این نوروترانسمیترهای اصلی، دهها نوروترانسمیتر فرعی هم هستند که به همراه نوروترانسمیترهای اصلی ترشح میشوند. برای هر نوروترانسمیتر، گیرندههای متعددی وجود دارد. هر گیرنده پاسخ مخصوص به خود را در برخورد با نوروترانسمیتر میدهد. به این ترتیب پاسخی که یک نورونِ پس سیناپسی به یک نوروترانسمیتر میدهد به تعداد و نوعِ گیرنده های موجود روی سطح آن بستگی خواهد داشت.
اجزاء بافت شناختی مغز و نخاع (شکلهای ۳، ۱۳، ۱۴، و ۱۵)
مادۀ خاکستری، عمدتاً از جسم سلولی نورونها تشکیل شده است همراه با تارهای عصبی. در مخ و مخچه مادۀ خاکستری عمدتاً در سطح قرار دارد و قشر (کورتکس) نامیده میشود. مادۀ سفید از تارهای عصبیِ نورونها تشکیل شده که مخ و مخچه عمقیتر قرار گرفته است. در همین مناطقِ عمقی نیز تجمعاتی از جسمهای سلولی (مادۀ خاکستری)، وجود دارد که به “هسته” موسوم هستند مانند هسته های ساقۀ مغز و هسته های تالاموس. در نخاع، مادۀ خاکستری در عمق و مادۀ سفید در سطح قرار گرفته است.
قسمتی از مادۀ سفید در دستگاه عصبی مرکزی که دارای تارهای عصیی موازی هستند، از جای معینی به هدف معینی گسیل شده اند و وظیفۀ کلانِ مشخصی دارند. به این دسته از تارهای عصبیِ موازی “ریس” میگوییم.
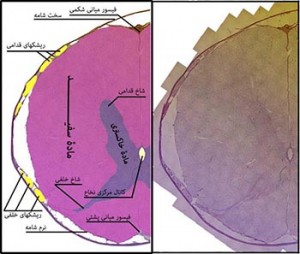 شکل ۳- نخاع؛ مادۀ خاکستری در نخاع در عمق، و مادۀ سفید در محیط قرار گرفته است. عکس سمت راست از به هم پیوستن چند ده عکس مجزا با استفاده از نرم افزار فوتوشاپ به دست آمده. قسمتهایی از این نخاع در بزرگنمایی بالاتر در شکل شمارۀ ۴ نمایش داده شده است. برای مشاهدۀ عکس بزرگتر و توضیحات بیشتر روی عکس کلیک کنید.
شکل ۳- نخاع؛ مادۀ خاکستری در نخاع در عمق، و مادۀ سفید در محیط قرار گرفته است. عکس سمت راست از به هم پیوستن چند ده عکس مجزا با استفاده از نرم افزار فوتوشاپ به دست آمده. قسمتهایی از این نخاع در بزرگنمایی بالاتر در شکل شمارۀ ۴ نمایش داده شده است. برای مشاهدۀ عکس بزرگتر و توضیحات بیشتر روی عکس کلیک کنید.
رنگامیزی هماتوکسیلین ائوزین، نخاع موش آزمایشگاهی سفید بزرگ، بزرگنمایی اصلی؛ ۱۰۰برابر. تهیه شده در کارگاه فرزاد. ۱۳۹۲
.
.
.
.
.
شکل ۴- جزئیات میکروسکوپی نخاع. برای مشاهدۀ عکس بزرگتر و توضیحات بیشتر روی عکس کلیک کنید.
سلولهای گلیال، از نظر شکل بسیار متنوعاند. هستۀ آنها بسیار کوچکتر از هستۀ نورونها است. خود سلولها نیز از نورونها عموماً کوچکتراند. هستکهای آنها نیز مشخص نیستند. سیتوپلاسم سلولهای گلیال اگر اصلاً قابل دیدن باشد، بسیار کم رنگ میگیرند. تشخیص انواع سلولهای گلیال از یکدیگر در رنگامیزیهای معمولی تقریباً ناممکن است.
سلولهای نوروگلیا یا گلیا، سلولهایی هستند که مستقیماً در کار انتقال پیام عصبی دخالت ندارند و به چند دسته تقسیم میشوند:
ستاره سلها؛ آستروسیتها یا آستروگلیا یا سلولهای ستاره ای؛ هم از جهت مکانیکی و هم از جهت متابولیکی، از نورونها حمایت می کنند. هنگامی که به دستگاه عصبی مرکزی آسیبی وارد شود و قسمتی از آن از بین برود، آستروسیتها در تشکیل بافت اسکارِ جایگزین، نقش مهمی دارند. استطاله های ستاره سلها از یک سو در ارتباط با عروق خونی هستند که به آنها، زائدۀ پایکیِ حولِ عروقی میگوییم.
|
گلیا در زبان یونانی، همخانوادۀ “گلو” یعنی چسب است. محققین متقدم، تنها وظیفه ای که به ذهنشان رسیده تا به این سلولهای درهم و برهم نسبت بدهند همین بوده است که مانند چسبی، نورونها، که کارکرد اصلیِ دستگاه عصبی را بر عهده دارند را در کنار هم نگاه دارد. هنگامی که رودولف ویرشو این کلمه را برای بافتی که نورونها در میان آن استقرار داشتند وضع میکرد، به این که این بافت تماماً از سلول تشکیل شده توجه زیادی نداشت و به کل آن نام “چسب عصبی” را داد [۱]. هنوز نیز کارکرد این سلولها تا حد زیادی ناشناخته است. تعداد آنها را تا همین چندی پیش یک تریلیون تخمین زده بوده اند [۶] اما تحقیقات اخیر تعداد آنها را بسیار کمتر از این دانسته است [۵]. اما محققین هر چه از تعداد آنها کم می کنند بر اهمیت آنها میافزایند. وجود گیرندههایی برای نوروترانسمیترها روی غشاء آنها دخالت آنها و احاطۀ تقریبا همۀ سیناپسها توسط سلولهای گلیا حضور فعال آنها را در انتقال پیام عصبی هر چه بیشتر تأیید میکند [۸] و[۹]. اخیراً دانشمندان تعدادی از آستروسیتهای انسانی را به مغز موش تزریق کرده اند و در کمال شگفتی دریافته اند که موشهای دریافت کنندۀ این آستروسیتها در یادگیری قویتر شده اند [۱۰]. در آزمایشاتی که روی مغز جسد برخی نوابغ انجام شده نیز ادعا شده که تعداد سلولهای گلیال مغز آنها با دیگران متفاوت است. به نظر میرسد، این سلولها چیزی بیش از یک چسب معمولی باشند که نورونها را فقط در کنار هم نگاه داشته اند. هنوز تا شناخت کارکرد دقیق آنها در این عمل، راهی طولانی برای محققین وجود دارد. |
الیگودنروسیتها؛ یا الیگوگلیا استطاله هایی کوتاهتر و کم تعدادتر دارند. این سلولها معادل سلولهای شوان هستند و در دستگاه عصبی مرکزی کار میلین سازی را بر عهده دارند. میلین یک لایه لیپیدی است که گرداگردِ آکسون را احاطه کرده و موجب سرعت انتقال بیشتر پیام عصبی در آن میشود. این روکش لیپیدی را در اعصاب محیطی، سلولی به نام سلول شوان میسازد و در مغز و نخاع ساختن این لایه بر عهدۀ الیگودندروسیت است. در اعصاب محیطی، یک سلول شوان متکفّلِ میلینه کردنِ قسمتی از فقط یک آکسون است اما در دستگاه عصبی مرکزی، یک الیگودندروسیت ممکن است چند قطعه از چند آکسون را میلینه کند (شکل ۵). در رنگامیزی معمولی هما.ئو. در نخاع، هسته هایی که در مادۀ سفید در یک صف قرار گرفته اند معمولاً الیگودندروسیت هستند.
میکروگلیا؛ سلول کوچکی است که بر خلاف نورونها و دیگر سلولهای گلیا منشاء جنینی متفاوتی دارد و تبار سلولیِ آن با تبار سلولیِ ماکروفاژها و مونوسیتها یکی است. کارکرد این سلولها نیز با بقیۀ سلولهای گلیایی متفاوت است و عملکرد آنها شبیه عملکرد ماکروفاژ است؛ هنگامی که به بافت عصبی مرکزی آسیبی وارد شود، این سلولها تکثیر یافته و تغییر شکل میدهند و به صورت سلولهای ریزهخوار عمل می کنند.
از بالا تا پایین نخاع، در محور مرکزیِ آن، یک لولۀ باریک قرار دارد که خالی از سلولهای پارانشیم نخاع است و با مایع مغزی-نخاعی پر شده است. در درونیترین مناطق مغز نیز فضاهای خالیی وجود دارد که با این قناتِ مرکزیِ نخاع مرتبط هستند و درون آنها نیز از مایع مغزی نخاعی آکنده است. در واقع مایع مغزی نخاعی در این فضاها در جریان و تبادل است. این فضای خالی از سلولهای عصبی و آکنده از مایع مغزی نخاعی را یک لایه سلول پوششی از درون، آستر کرده است که به سلولهای آن سلولهای آپاندیمی میگویند. این سلولها بر خلاف اغلب سلولهای پوششی فاقد اتصال تنگ هستند. به این ترتیب اگر چه شبیه به بافت پیوندی هستند و مرزی را میان مایع مغزی نخاعی از یک سو، و پارانشیم مغز و نخاع از سوی دیگر میسازند اما یک مرز تمام عیار و نفوذناپذیر را نمیسازند. نبود اتصال تنگ، تبادل مایعات و مواد از مایع مغزی نخاعی به داخل مغز و نخاع از طریق این آسترِ پوششی را ممکن میسازد. همین سلولها در بعضی از مناطق مغز که سد مغزی نخاعی، ناقص است، به سلولهای مژکداری تبدیل میشوند که ورود و خروج مواد را به داخل پارانشیم مغز مدیریت می کنند. لازمۀ مدیریت و کنترل ورود و خروج مواد از بافت پوششی وجود اتصال تنگ است و سلولهای آپاندیمی در این مناطقِ خاص به این اتصالات مجهز هستند. به این سلولهای آپاندیمی در این مناطق خاص، تانی سیت هم میگویند.
مقصود از دستگاه عصبی محیطی هر بافت عصبی خارج از مغز و نخاع است. هنگامی که از دستگاه عصبی محیطی صحبت میشود آن چه بیشتر جلب توجه میکند، خودِ اعصاب محیطی هستند، در حالی که اعصاب، تنها یکی از سه جزء اصلی دستگاه عصبی محیطی هستند. در کنار اعصاب، عقده های عصبی و پلکسوسهای عصبی نیز اجزاء بسیار مهم این دستگاه هستند. نوروترانسمیترهای اصلی دستگاه عصبی محیطی استیل کولین و نورادرنالین هستند.
اعصاب محیطی
عصب محیطی عبارت است از یک دسته از تارهای عصبی که وظیفۀ انتقال پیام عصبی را از بدن به مغز و نخاع یا از مغز و نخاع به بدن بر عهده دارد. اعصاب عموماً فاقد جسم سلولی هستند و فقط شامل آکسونها هستند. یک عصب شبیه یک کابل تلفن است که فقط پیام را به/از مرکز مخابرات منتقل میکند. سیمهای مسی داخل کابل تلفن، در حال آوردن یا بردنِ پیام به یا از مرکز تلفن هستند. به تارهای عصبی که پیامی را به نخاع و مغز میبرند، تارهای آوران و به تارهایی که پیام عصبی را از مغز یا نخاع به اندامها منتقل می کنند، تارهای عصبیِ وابران میگوییم. تارهای عصبی وابران را نورونِ حرکتی نیز میخوانند؛ این نورونها به تارهای عضلانی یا به غدد ختم میشوند. برخی از مؤلفین همۀ نورونهای آوران را نورونِ حسی مینامند، بهتر است میان نورونهای حسی که پیامهای حسی را به مرکز منتقل می کنند و دیگر نورونهای آوران تمایز قایل شد. برخی از نورونهای آوران پیامهایی را از عضلات به مرکز منتقل می کنند که در تعادل اندامهای حرکتی دخالت دارند. بسیاری از مؤلفین ترجیح میدهند که به این دسته از نورونها، نورون حسی اطلاق نکنند. عصبی که صرفاً حاوی تارهای حسی یا حرکتی باشد تقریباً در بدن انسان وجود ندارد. بیشتر اعصاب محیطی، اعصاب مخلوط هستند به این معنی که حاوی تارهای آوران و وابران با هم هستند.
نادر بودنِ عصبهای خالص حرکتی یا عصبهای خالص حسی، عجیب نیست. آیا میتوان مثلا محله یا منطقه ای را فرض کرد که کابلهای تلفن، به صورت یکطرفه فقط پیام ها را ببرند یا فقط آنها را به مرکز برسانند؟ اگر عصبی برای مثال انگشتان دست راست را تعصیب میکند بهتر است که عصبی که مانند یک کابل پر از سیم، به آنجا میرود هم حاوی تارهای حسی باشد که از پوستِ دست به مرکز پیام برساند و هم حاوی تارهای حرکتی باشد که دستورات انقباض و انبساط را به عضلات انگشتان میبرد.
فرض کنید که درداخل یک کابل تلفن فقط و فقط سیمهای مسی، بدونِ روکشهای پلاستیکی وجود داشته باشد. چنین کابلی در انتقال پیام میان مراکز و اندامها کاملاً ناکارا خواهد بود، چرا که پیامها در میان راه از سیمی به سیم دیگر منحرف خواهد شد. روکشهای پلاستیکی سیمهای مسی، از فرسایش و خوردگی آنها نیز جلوگیری می کنند و از نظر مکانیکی نیز از آنها حمایت می کنند. آکسونهای داخل یک عصب نیز از چنین روکشهایی برخوردارند و ساختارهای متنوعی در عصب وجود دارند که از آنها حمایت مینمایند. میلین و اندونوریوم از جملۀ این ساختارها هستند که متعاقباً با آنها آشنا خواهید شد.
آکسونِ میلینه و آکسونِ غیر میلینه (شکل ۵)
همانطور که قبلاً اشاره شد، یک آکسون ممکن است تا چند سانتیمتر و حتی تا یک متر درازا داشته باشد. به این ترتیب محتوای این آکسون و غشاءِ آن، بسیار دور از هستۀ سلول است و ادارۀ اعمالِ حیاتیِ این آکسونِ دور از هسته باید به طریقی، تأمین شود. بسیاری از نیازهای حیاتیِ این قطعاتِ دور از هسته بر عهدۀ سلول شوان است. به این ترتیب که سلول شوان، که در لابهلای آکسونها قرار گرفته اند، سیتوپلاسم خود را کِش داده، دورِ آکسونها را احاطه میکند و به این ترتیب علاوه بر عایق کردنِ آکسون برخی از نیازهای متابولیک آن قطعه از آکسون را که احاطه کرده است، تأمین میکند. یک سلول شوان ممکن است قسمتی از سیتوپلاسم خود را دور چند صد میکرون از این آکسون محیط کند و در عین حال ممکن است این عمل را با ۳۰ آکسون دیگر نیز انجام دهد. به چنین آکسونی، آکسون غیر میلینه میگویند. در آکسونِ غیر میلینه، آن فضایی که غشاء و سیتوپلاسمِ شوان در اختیار آکسون قرار داده تا آکسون در آن جایگیر شود را مزاکسون میخوانند.
گاهی یک سلول شوان همۀ سیتوپلاسم خود را فقط به یک تکه از یک آکسون اختصاص میدهد و در این حالت سیتوپلاسم خود را به همراهِ غشاءِ خود دور این تکه از آکسون دهها بار میپیچاند. البته در این حالت در این ناحیه، چیز زیادی از سیتوپلاسم باقی نمیماند، و تنها چند صد لایه غشاء سلول است که دور تا دور این تکه از آکسون را احاطه کرده است. به چنین آکسونی، آکسونِ میلینه میگویند. به این ترتیب ممکن است یک آکسون میلینه در طولِ مسیر خود چند صد یا چند هزار سلول شوان را به خدمت گرفته باشد؛ اما توجه داریم که هر شوان، تنها یک تکه از یک آکسون را با غشاء در هم پیچیدۀ خود احاطه کرده است. [۱۱]
همچنین باید توجه داشته باشیم که آکسون غیر میلینه نیز در طول مسیر خود شوانهای متعدد را به خدمت گرفته اما اولاً شوان فقط به سادگی قسمتی از آن را احاطه کرده و دور آن نپیچیده است و ثانیاً ممکن است این آکسون یکی از دهها آکسونی باشد که این شوانِ خاص آن را حمایت کرده است.
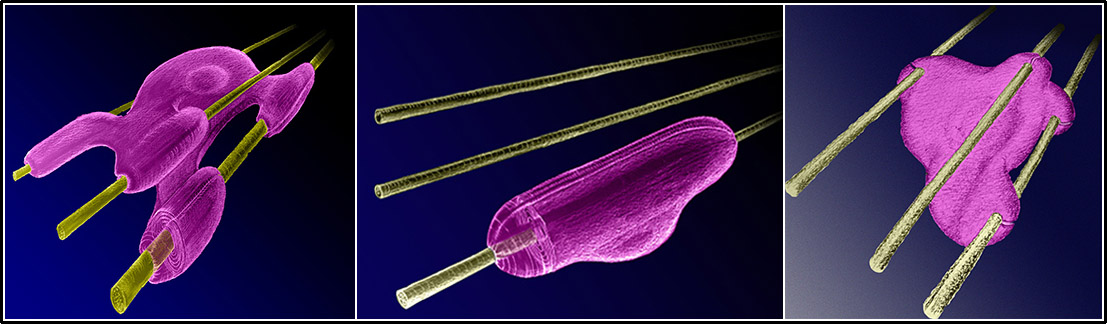
شکل ۵- آکسون میلینه و غیر میلینه؛ شکل شماتیکِ سمت راست، چند آکسون غیر میلینه را نشان میدهد که توسط سلول شوان (یا الیگودندروسیت) حمایت شده است. هر سلول حامی، قطعاتی از چندین آکسون مجاور را احاطه میکند. سیتوپلاسمِ سلول شوان همۀ محیط آکسون را به طور کامل احاطه نمیکند، شکاف بسیار بسیار باریکی که دو قسمت سیتوپلاسم شوان در آن شکاف به هم نزدیک میشوند مدخل مزاکسون است. شکل میانی و شکل سمت چپ، آکسون میلینه را در اعصاب محیطی و دستگاه عصبی مرکزی نشان میدهند. شوان در اعصاب محیطی و الیگودندروسیت در دستگاه عصبی مرکزی با گرداندن چند صد بارۀ سیتوپلاسم خود به گرد آکسون آن را میلینه میکند. هر شوان فقط یک قطعه از یک آکسون را فرا میگیرد، اما هر الیگودندروسیت ممکن است چند قطعه از چند آکسون را حمایت کند. در آکسونهای میلینه نیز مزاکسون تعریف شده است. در این طرحهای شماتیک، مقیاسها رعایت نشده است.
در آکسونِ میلینه همانطور که گفتیم؛ یک شوان تنها قسمتی از مسیر آن را با لایه های متعددی از غشاءِ خود احاطه میکند. و قطعۀ مجاور، در اختیار شوانِ دیگری قرار میگیرد تا آن را میلینه کند. در فاصلۀ دو روکشِ میلینی فاصلۀ بسیار کوچکی وجود دارد که برهنه است و میلین آن را نپوشانده است. این فاصلۀ کوچک را به افتخار کاشفِ آن، گرۀ رانویه نام گذاری کرده اند. به این ترتیب درطولِ یک آکسونِ میلینه، به فواصلِ معین، گرۀ رانویه وجود دارد (شکل۷). این ساختار، در مجموع موجب میشود که سرعتِ جریانِ پیام عصبی در آکسون به مراتب سریعتر شود. برای فهم کیفیت این عمل و اثر این ساختارِ تخصصی در سرعت جریانِ پیام لازم است فیزیولوژیِ جریان عصبی و پتانسیل عمل را بشناسیم که در منابع فیزیولوژی به تفصیل در این باره بحث شده است. یکی از بهترین منابعی که این کارکرد به همراه شیوۀ انتقال پیام عصبی از آغاز تا پایان در آن شرح داده شده است کتاب اصول علوم عصبی به سرویراستاریِ “کاندل” است [۶].
تارهای عصبی موجود در اعصاب را با توجه به ساختار و کارکردشان به سه نوع کلی تقسیمبندی می کنند:
تارهای نوع الف (A) تارهای میلینه هستند که ضخامتی در حدود ۴ تا ۲۰ میکرون دارند و پیام عصبی را با سرعتی بالا در حدود ۱۵ تا ۱۲۰ متر در ثانیه، انتقال میدهند. تارهای حرکتیِ محرّکِ تارهای عضلانیِ اسکلتی، و تارهای حسی از این نوع اند.
تارهای نوع ب (B) نیز میلینه هستند که قطری در حدود ۱ تا ۴ میکرون دارند و سرعت پیام در آنها حدود ۳ تا ۱۴ متر در ثانیه است. تارهای عصبی خودکارِ پیش عقده ای (فیبرهای عصبیِ اتونومِ پیش گانگلیونی) از این نوع هستند.
تارهای نوع ج (C) قطری کمتر از ۱ میکرون دارند و سرعت پیام در آنها از ۲ دهم تا ۲ متر در ثانیه است. تارهای خودکار و تارهای منتقل کنندۀ پیام درد، از این نوع هستند.
بافت پیوندی در عصب
دورادور هر آکسون را یک لایۀ نازک از بافت همبند پوشانده است و مانند تونلی در همۀ مسیر آکسون را همراهی میکند. این بافت حاوی صفحۀ پایه (لامینا بازال) و ملکولهای مربوط به این ساختار است. برخی ملکولهای موجود در صفحۀ پایه در آزمایشات، خاصیتهای مشخصی را از خود بروز داده اند که نشان دهندۀ آن است که برای ترمیم عصب و راهنماییِ عصب در مسیری مشخص در هنگام ترمیم و رشد، نقش بنیادین دارند. به همین دلیل است که بسیاری از محققین حوزۀ ترمیم اعصاب علاقمند به ملکولها و هندسۀ این ساختارها هستند. این لایۀ بسیار نازک را که تنها یک آکسون را احاطه کرده است، آندونوریوم نامیده اند. کلّ عصب نیز توسط یک لایۀ ضخیم از بافت پیوندی موسوم به اپینوریوم روکش شده است که نقشِ تمام عیاری در حمایت مکانیکیِ عصب دارد و آن را در مقابل نیروهای مکانیکی که به آن وارد میشود محافظت میکند. پِرینوریوم، لایه ای از بافت پیوندی است که از لحاظ ضخامت ما بین آندونوریوم و اپینوریوم است و در درون عصب یک دسته از آکسونها را در بر گرفته است. تفاوت پرینوریوم و آندونوریوم و نیز تفاوت آن با اپینوریوم تنها در ضخامت و ساختار ظاهری نیست، تراوایی و خاصیت نفوذپذیری پِرینوریوم با دو نظیر پیوندی خود متفاوت است. پرینوریوم نقشی مهم در ایجاد سد خونی-عصبی دارد و در ادامۀ مسیر خود به سد خونی – مغزی در نخاع و مغز میپیوندد.
به مجموعۀ آکسونهایی که درونِ یک پرینوریوم قرار گرفته اند، یک دسته (فاسیکول یا باندل) میگویند.
پرینوریوم همچنین مورد توجه جراحان نیز هست، از این جهت که ساختاری است که به جهت ضخامتِ قابل قبول آن میتوان آن را در صورت آسیب دیدن و بریده شدن، مورد جراحی قرار داد و به تکۀ مقابل خود بخیه زد؛ البته با نخهای بسیار ظریف و زیر میکروسکوپ. با دوختنِ پرینوریوم به جای دوختنِ صِرفِ اپینوریوم، نتیجۀ بهتری در ترمیم عصب به دست میآید.
در بافت همبندِ درونِ عصب، فیبروبلاست، ماکروفاژ و ماست سِل های متعدد وجود دارند. همچنین، بافت همبند عصب بسترِ عبورِ عروقی است که مثل هر جای دیگر برای انتقال اکسیژن و مواد غذایی به هر بافت زنده از جمله آکسونها ضروری هستند.
یک شریان همچنان که از اپینوریوم به داخل عصب نفوذ میکند، منشعب شده، باریک تر میشود و در پرینوریوم به شریانک (آرتریول) و در اندونوریوم به مویرگ تبدیل میشود و برای خدمت رسانی به آکسونها و سلولهای دیگر موجود در عصب آماده میشود.

.
.
.
.
شکل ۶- مقطع طولی عصب محیطی شکل ۷- گره رانویه شکل ۸- مقطع عرضی عصب محیطی


.
.
.
.
شکل ۹- مقطع عرضی عصب محیطی شکل ۱۰- مقطع طولی آکسون شکل ۱۱- مقاطع عرضی آکسونها
.
گانگلیون یا عقدۀ عصبی، تجمعی از سلولهای عصبی (جسم سلولی) خارج از دستگاه عصبی مرکزی هستند. مهمترین عقده های عصبی، عبارتاند از عقدۀ ریشۀ خلفی نخاعی و عقدۀ خودکار. گانگلیونهای اوتونوم (خودکار) یا در محاذاتِ ستون فقرات قرار گرفته اند یا در خود بافت در دیوارۀ دستگاه گوارش و مثانه (گانگلیون اینترادورال یا درون مرزی).
گانگلیونهای ریشۀ خلفی، حاویِ جسمِ سلولیِ آکسونهای آوران حسی هستند. این نورونها شکلی منحصر به فرد دارند و به آنها نورونهای شبهِ تک قطبی یا تک قطبیِ کاذب میگویند. نورونِ حسیِ گانگلیونِ خلفی در واقع یک آکسونِ ضخیم و طولانی است که پیام حسی را یکسره در طول مسیرِ خود به نخاع منتقل میکند. این آکسون در مسیر خود به سوی نخاع، در محل گانگلیون ریشۀ خلفی یک جسم سلولی دارد که نقش آن در تغذیۀ این آکسون است و نه تنها هیچ سیناپسی ندارد بلکه حتی پیام عصبیِ آکسونِ آن لازم نیست از آن رد شود و موقعیت آکسون به گونه ای است که، پیام عصبی میتواند بدون عبور از جسم سلولی، راه خود را به سوی نخاع پی گیرد. در واقع نورونِ شبهِ تک قطبی، دندریتی ندارد و فقط به یک آکسون متصل است. ستاره سلها دور تا دور هر جسم سلولی را احاطه کرده اند. و کلِّ گانگلیون توسطِ یک کپسول همبند احاطه شده است.
نورونهای موجود در گانگلیونِ عصبیِ خودکار، در مقابل؛ مانند اغلب نورونهای دیگر، دارای آکسون و دندریت هستند و بی استتثناء سیناپسهایی دارند که پذیرای پیام عصبی است.
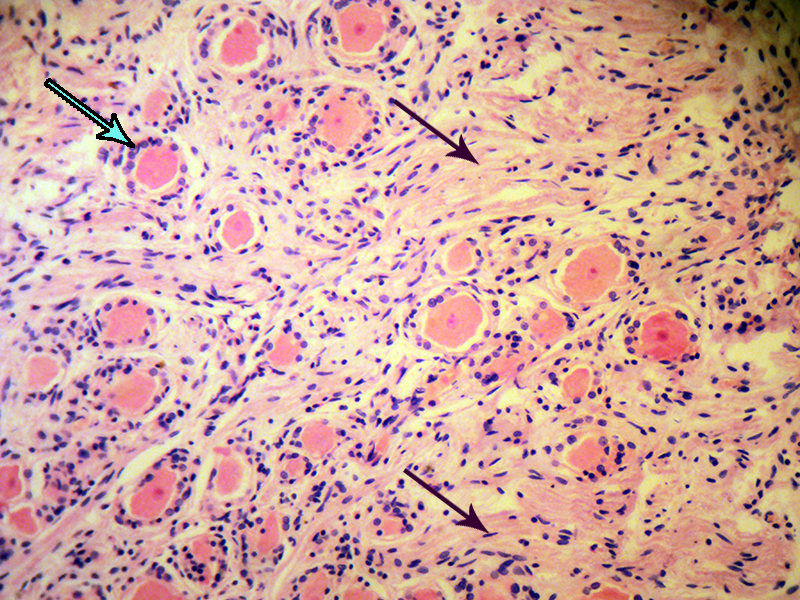 شکل ۱۲-الف – منظرۀ عمومی گانگلیون خلفی، جسم سلولی این نورونها بسیار بزرگ است و هستۀ بسیار کمرنگ دارند. در اطراف هر جسم سلولی چندین ستارهسل با هستههای کوچکترِ پر رنگ دیده میشود. اجسام نیسل به سیتوپلاسمِ حیاط هسته منظرۀ دانه دانه داده است. دیدن آکسونی که از جسم سلولی جدا شده و از میان ستاره سلها بیرون آمده، در مقاطعی با ضخامت کمتر از ۶ میکرون، بختِ بلندی میخواهد. این مقطع از گانگلیون حسی عصب گاسر یا عصب سه قلو تهیه انسان تهیه شده است که بافت آن با بافت گانگلیون شاخ خلفی نخاعی بسیار شباهت دارد. از لابه لای نورونها، دسته های آکسونها درکنار هم طی به محیط یا به مرکز طی مسیر میکنند (پیکان سیاه). پیکان آبی، به مجموعهای از ستاره سلها نشانه رفته است که یک جسم سلولی را احاطه کرده اند.بزرگنمایی اصلی: ۱۰۰ برابر؛ رنگامیزی هماتوکسیلین و ائوزین – گانگلیون گاسر انسانی – تهیه شده در کارگاه فرزاد – ۱۳۷۶
شکل ۱۲-الف – منظرۀ عمومی گانگلیون خلفی، جسم سلولی این نورونها بسیار بزرگ است و هستۀ بسیار کمرنگ دارند. در اطراف هر جسم سلولی چندین ستارهسل با هستههای کوچکترِ پر رنگ دیده میشود. اجسام نیسل به سیتوپلاسمِ حیاط هسته منظرۀ دانه دانه داده است. دیدن آکسونی که از جسم سلولی جدا شده و از میان ستاره سلها بیرون آمده، در مقاطعی با ضخامت کمتر از ۶ میکرون، بختِ بلندی میخواهد. این مقطع از گانگلیون حسی عصب گاسر یا عصب سه قلو تهیه انسان تهیه شده است که بافت آن با بافت گانگلیون شاخ خلفی نخاعی بسیار شباهت دارد. از لابه لای نورونها، دسته های آکسونها درکنار هم طی به محیط یا به مرکز طی مسیر میکنند (پیکان سیاه). پیکان آبی، به مجموعهای از ستاره سلها نشانه رفته است که یک جسم سلولی را احاطه کرده اند.بزرگنمایی اصلی: ۱۰۰ برابر؛ رنگامیزی هماتوکسیلین و ائوزین – گانگلیون گاسر انسانی – تهیه شده در کارگاه فرزاد – ۱۳۷۶
.
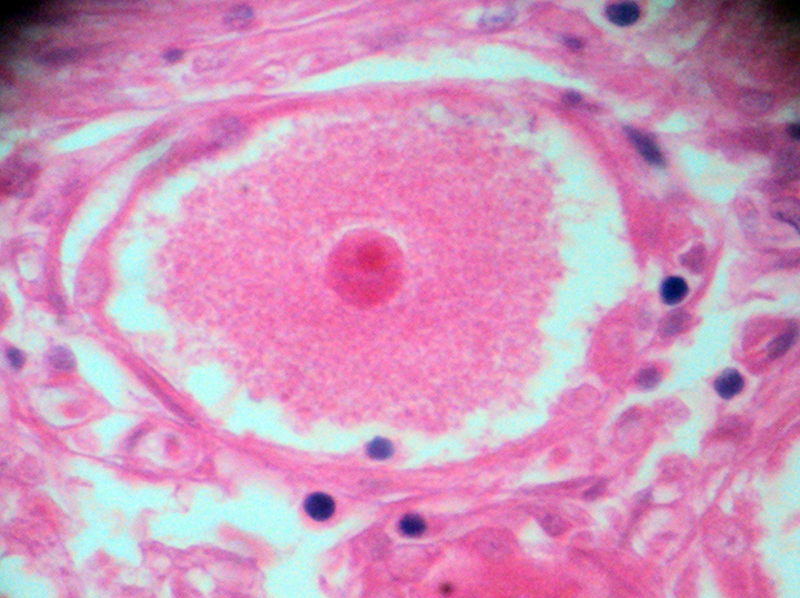
شکل ۱۲-ب – جسم سلولی نورون گانگلیون حسی؛ هسته و هستک در وسط و سیتوپلاسم دانه دانه در اطراف آن. این دانهها را اجسام نیسل میخوانند و در واقع، شبکۀ آندوپلاسمیک است به همراه ریبوزومهای آزاد یا ریبوزومهای چسبیده به این شبکه. به تفاوت رنگ و اندازۀ هستۀ این نورون با هستۀ ستارهسلهای اطرافِ آن توجه کنید.بزرگنمایی اصلی: ۴۰۰ برابر؛ رنگامیزی هماتوکسیلین و ائوزین – گانگلیون گاسر انسانی – تهیه شده در کارگاه فرزاد – ۱۳۷۶
.
.
.
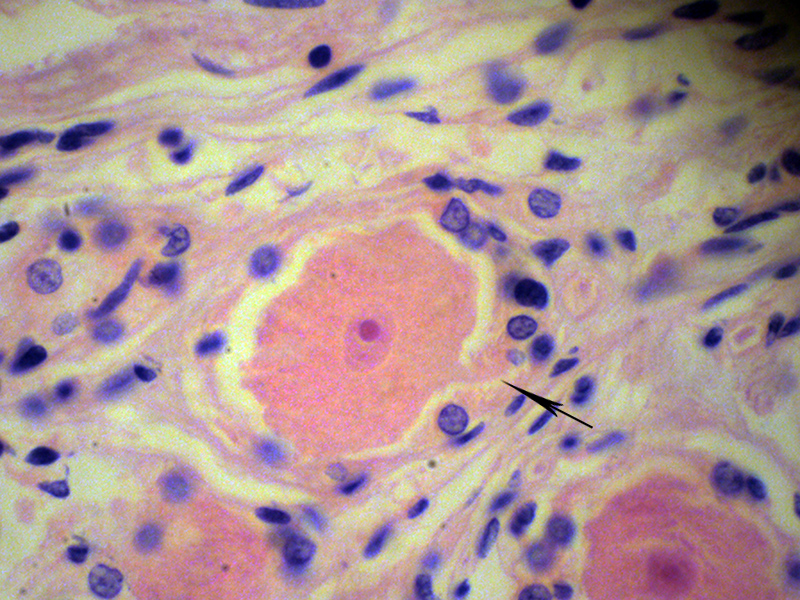
شکل ۱۲-ج- جسم سلولی نورون گانگلیون حسی؛ هسته و هستک در وسط و سیتوپلاسم دانه دانه در اطراف آن. از سمت راستِ حیاطِ هسته، استطالهای بیرون زده (پیکان سیاه) و از میان ستارهسلها خارج شده که به احتمال بسیار زیاد، آکسون است.بزرگنمایی اصلی: ۴۰۰ برابر؛ رنگامیزی هماتوکسیلین و ائوزین – گانگلیون گاسر انسانی – تهیه شده در کارگاه فرزاد – ۱۳۷۶
.
.
.
.
.
.
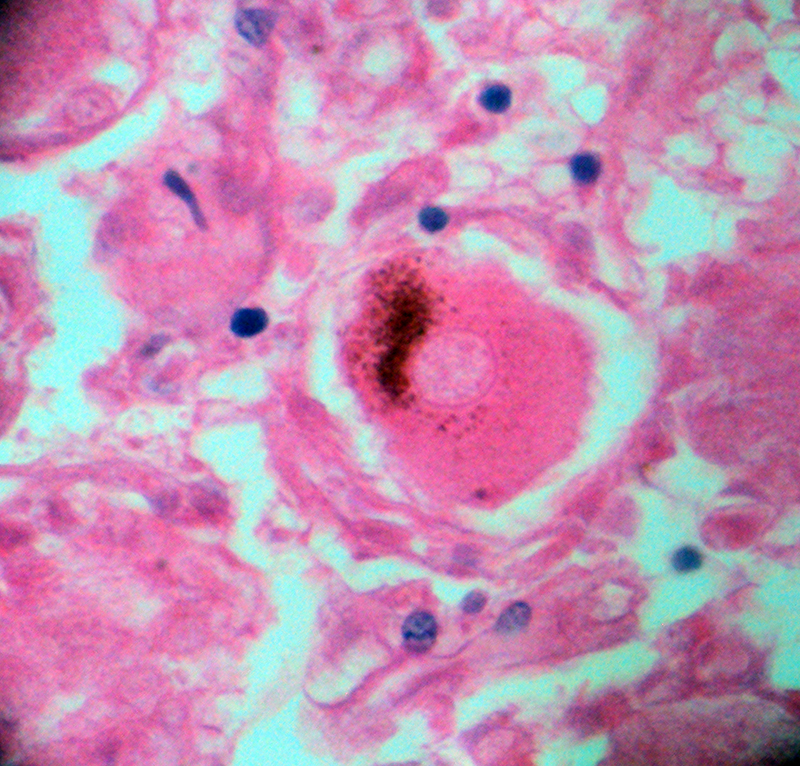
شکل ۱۲-د – جسم سلولی نورون گانگلیون حسی؛ در سیتوپلاسم این سلول دانه های قهوه ای رنگ دیده میشود که لیپوفوشین است. وجود لیپوفوشین در حدی معین، در برخی از سلولهای افراد مسن طبیعی است. برای مشاهدۀ عکس بزرگتر ، روی آن کلیک کنید.بزرگنمایی اصلی: ۴۰۰ برابر؛ رنگامیزی هماتوکسیلین و ائوزین – گانگلیون گاسر انسانی – تهیه شده در کارگاه فرزاد – ۱۳۷۶
.
.
.
.
.
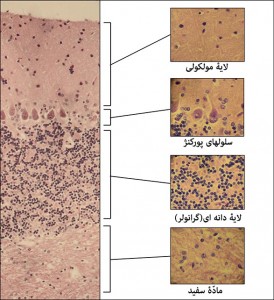
شکل ۱۳- مخچه و لایه های آن؛ لایه های مخچه از سطح به عمق عبارتند از لایۀ مولکولی، لایۀ سلولِ پورکنژ و لایۀ گرانولر. در زیر این مجموعه جسم سفید قرار گرفته. برای مشاهدۀ عکس بزرگ و توضیحات بیشتر روی آن کلیک کنید.رنگامیزی هماتوکسیلین و ائوزین – مخچۀ موش آزمایشگاهی سفید بزرگ – تهیه شده در کارگاه فرزاد – ۱۳۹۲
.
.
.
.
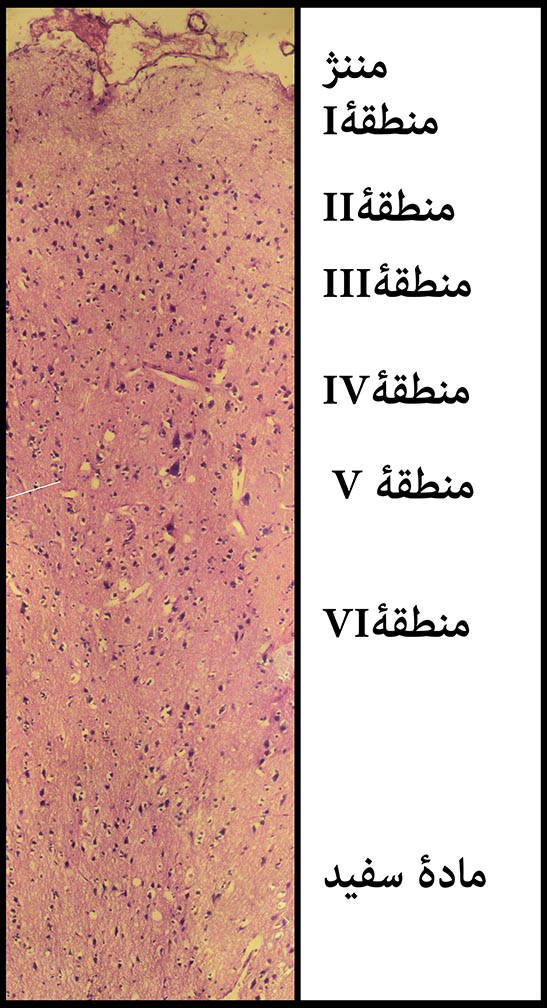
شکل ۱۴- مخ و لایه های ششگانۀ قشر (کورتکس) آن؛ بر خلاف نخاع مادۀ خاکستری در مغز در سطح و مادۀ خاکستری در عمق قرار گرفته است. از سطح تا عمقِ کورتکس که به حدّ مادۀ سفید میرسد، را از لحاظ ترکیب و نوع سلولها به ۶ منطقه تقسیم کرده اند. نمیتوان میان این مناطق، خطِّ مشخصی کشید و این فقط یک مرزبندی نسبی است. سلولهای هرمی درمنطقۀ ۳ و سلولهای هرمی غولپیکر در منطقۀ ۵ قرار دارند. برای مشاهدۀ عکس بزرگتر، روی آن کلیک کنید. با عکس ۱۵ مقایسه کنید. بزرگنمایی اصلی؛ ۱۰۰ برابر. رنگامیزی هماتوکسیلین و ائوزین – مخ موش آزمایشگاهی سفید بزرگ – تهیه شده در کارگاه فرزاد – ۱۳۹۲
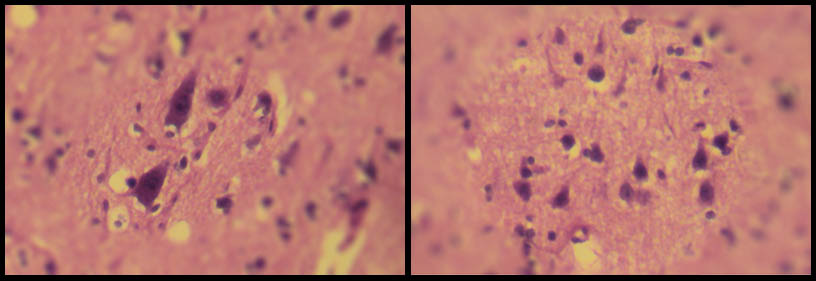 شکل ۱۵- سلولهای هرمی در مخ؛ سلولهای هرمی (درمنطقۀ ۳ مخ) در عکس سمت راست و سلولهای هرمی غولپیکر (در منطقۀ ۴ مخ) در عکس چپ دیده میشود. این دو عکس قسمتهایی از عکس شکل ۱۴ هستند. بزرگنمایی اصلی؛ ۱۰۰ برابر. رنگامیزی هماتوکسیلین و ائوزین – مخ موش آزمایشگاهی سفید بزرگ – تهیه شده در کارگاه فرزاد – ۱۳۹۲
شکل ۱۵- سلولهای هرمی در مخ؛ سلولهای هرمی (درمنطقۀ ۳ مخ) در عکس سمت راست و سلولهای هرمی غولپیکر (در منطقۀ ۴ مخ) در عکس چپ دیده میشود. این دو عکس قسمتهایی از عکس شکل ۱۴ هستند. بزرگنمایی اصلی؛ ۱۰۰ برابر. رنگامیزی هماتوکسیلین و ائوزین – مخ موش آزمایشگاهی سفید بزرگ – تهیه شده در کارگاه فرزاد – ۱۳۹۲
پانوشت:
[۱] بر خلاف انتظار نوروگلیا از لحاظ دستوری جمع نیست است و اگر شبیه به کلمات جمع در زبان لاتین به “ا” ختم میشود اما متشکل است از نورون به معنی عصب و گلیا به معنی چسب. در واقع این کلمه اساسا در زبان انگلیسی شکل مفرد ندارد. برای نقش ویرشو در کشف و نامگذاریِ نوروگلیا مراجعه کنید به:
برای وضعیت دستوری کلمه گلیا در زبان انگلیسی مراجعه کنید به:
http://oxfordindex.oup.com/view/10.1093/oi/authority.20110803100230112
[۲]
و
http://www.translatingtime.net/home
[۳]
[۴] برای دیدن عکسی که از یکی از تیغه های تهیه شده توسط سانتیاگو رامون کاخال تهیه شده به
http://www.scholarpedia.org/article/File:Pyramidal_cell_Golgi_method.png
{kind=link}
و برای دیدن طراحی او از آنچه دیده به
http://en.wikipedia.org/wiki/File:PurkinjeCell.jpg
{kind=link}
مراجعه کنید. در این طراحی او چند نورون چند قطبی را نمایانده است. کاخال به خوبی خارها را تشخیص داده است و دندریتهای زبر را ترسیم کرده است.
برای دیدن نقاشی کاخال از نورونهای دو قطبی شبکیۀ چشم به
http://commons.wikimedia.org/wiki/File:Cajal_Retina.jpg
{kind=link}
مراجعه کنید.
[۵] سالهاست که تعداد نورونهای مغز انسان را در حدود ۱۰۰ میلیارد، و تعداد سلولهای گلیال را ۱۰ برابر این تعداد تخمین زده اند. در آخرین تحقیقی که در این باره انجام شده، تعداد نورونهای مغز را حدود ۹۰ میلیارد و تعداد سلولهای گلیال را در همین حدود دانسته اند. برای دیدن این تحقیق و شیوۀ بدیعش در شمارش سلولهای مغزی، مراجعه کنید به مقالۀ
[۶] در بارۀ تعداد و انواع نورونها و اتصالات آنها با هم مراجعه کنید به کتاب اصول علوم اعصاب، چاپ پنجم، فصل دوم
Kandel ER, 2012. Principles of Neural Science, 5th ed. McGraw-Hill, New York
در چاپ چهارم همین کتاب که در سال ۲۰۰۰ میلادی به چاپ رسید تعداد سلولهای گلیال ۱۰ برابر نورونها در مغز گفته شده است.
پتانسیل عمل در این کتاب در فصول مختلف به خوبی شرح داده شده است.
[۷] برای مشاهدۀ تصاویری از سیناپس با میکروسکوپ الکترونی به اینجا
http://www.uni-mainz.de/FB/Medizin/Anatomie/workshop/EM/EMSynapseE.html
مراجعه کنید. در این تصاویر سیناپس متقارن و نامتقارن را میتوانید بیابید.
[۸] نشانه هایی که نقشهای متنوع سلولهای گلیال را در انتقال پیام عصبی تأیید میکند در این مقاله دوره شده است؛
[۹] تأثیر و نقش ستاره سلها در تنفس موضوع این تحقیق است؛
[۱۰] پیوند ستاره سلهای انسانی به موش؛
[۱۱] پویانما؛ میلینه شدنِ آکسون توسطِ شوانسل
http://www.lab.anhb.uwa.edu.au/mb140/CorePages/Nervous/Nervous.htm#NERVES
دندریت یعنی چی ؟؟؟؟؟؟؟؟
در لغت یعنی شاخه درخت اما در بافت شناسی اعصاب همان است که در متن گفته شده است.
بافت چندقطبی یایک قطبی یعنی چی
چیزی به نام “بافت چند قطبی” را نشنیده و ندیده ام.
سلام مطالب را می شود به صورت pdf به ایمیل ارسال بفرمائید تشکر عالی بود