
بافت شناسی نظری > بافت عضلانی
در این صفحه مییابید:
دو واژۀ “حرکت” و “زندگی” در بسیاری از موارد مترادف تلقی میشوند. هیچ سلولِ بیحرکتی در دنیا پیدا نمیشود. حرکت، منحصر به پروازِ شاهین با سرعت نزدیک به ۴۰۰ کیلومتر بر ساعت [۱] یا مهاجرت ۸۰ هزار کیلومتری پرستوی قطبی [۲]نیست. مهاجرتهای بسیار کند و کم مسافتِ سلولهای ابتداییِ جنینی نیز حرکت است و از لوازمِ ادامۀ حیات. همچنین است حرکتهای منظمِ میکروسکوپی اندامکهای داخل سلولی است که تقسیم سلولی و تکثیر موجودات زنده را امکان پذیر میکند.
حرکت سلول در محیط کشت  حرکت اندامکها در تقسیم سلولی
حرکت اندامکها در تقسیم سلولی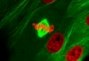
هر جا حیات هست، حرکت هست؛ با کمی مسامحه نیز میتوان گفت که هر جا حرکتِ حیاتی هست، اکتین و میوزین یا موتورهای ملکولی دیگر نیز هست. همان میوزین و اکتینی که موجب حرکت آمیبی در آغازیان و تغییر شکل غشاء در قارچهاست، عامل حرکتِ بازو و انگشتانِ ما انسانهاست.
در واقع اکتین و میوزین فقط عامل حرکت عضلات نیستند بلکه این دو ملکول در سلولهای غیر عضلانی نیز وجود دارند و در همین سلولها کارکردهای متنوع و از جمله ایجاد حرکت دارند.
برای دیدن حرکتِ اندامکهای داخل سلول و حرکتِ سریع یک تکسلولی به این صفحه مراجعه کنید.
بهترین راه فهمیدنِ بافت شناسی و فیزیولوژیِ عضله، این است که اول با میوزین یا موتورهای پروتئینی و عملکردِ این ملکولها در سادهترین موجودات آشنا شویم و سپس به بافت شناسی عضلات صاف و مخططِ اسکلتی و قلبی بپردازیم.
میوزین در همۀ یوکاریوتها جز جلبکهای قهوهای و برخی از آغازیان وجود دارد [۳] – (Syamaladevi-2012).
میوزین یک پروتئین سنگین است که از سه ناحیه که معمولاً به سر، بازو (یا گردن) و دم نامگذاری میشوند؛ تشکیل شده است. اساسِ حرکتِ عضله، تحرک ناحیۀ بازو و قدرتِ چسبندگیِ ناحیۀ سر به اکتین یا پروتئینهای دیگر است.
مراحلِ حرکتِ میوزین روی رشتۀ اکتین به این ترتیب است:
۱- در حضور کلسیم، تروپومیوزین که دورِ رشتۀ اکتین تابیده شده است، جابجا شده، قسمتهایی از اکتین که به سَرِ میوزین چسبندگی دارند، نمایان میشوند.
۲- سرِ میوزین به اکتین میچسبد.
۳- در حضور منیزیم، آ.ت.پ. که به سرِ میوزین چسبیده، انرژی خود را رها میکند و از آن جدا میشود؛ این انرژی صرف خم شدنِ بازوی میوزین میشود.
۴- با وجود آ.ت.پ. کافی در محیط، یک آ.ت.پ به سر میچسبد و موجب جدا شدنِ سر از اکتین میگردد.
۵- بازو به حالت باز قرار میگیرد.
وجودِ کلسیمِ کافی در دسترسِ تروپونین، (که به تروپومیوزین اتصال دارد) موجب میشود که چرخۀ فوق تکرار شود.
چند پویانما در بارۀ حرکت میوزین و اکتین
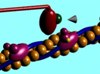


در سلولهای غیر عضلانی، میکروتوبول و اکتین، مانند ریل عمل میکنند که موتورهای پروتئینی روی آن حرکت میکنند. این موتورها ممکن است دو سر و دو بازو داشته باشند. وصل و قطع شدنِ مکررِ سر و خم و راست شدنِ بازوی این موتورهای پروتئینی، موجب حرکت میشود. دُمِ آنها نیز، به یک “کوله” متصل میشود که غرض از همۀ این سازکارها حمل همین کوله است.
حرکت موتورِ پروتئینی، در صورتی است که ریلِ زیرِ پای آنها، ثابت باشد. اما اگر میوزینها از ناحیۀ دم به جایی ثابت و سنگین متصل باشند. نیروی حاصل از خم شدنِ بازوی میوزین، موجب خواهد شد که ریل، و آن چه به ریل متصل است به سمت آنها کشیده شود.
اکتین در عضلۀ صاف از نوعِ اکتینِ رشتهای (f-actin) است. مونومرهای اکتین به هم میپیوندد و به صورت رشتههای دو تای در هم تنیدۀ بسیار درازی در میآید که از این سو به آن سوی سلول کشیده شدهاند. در واقع قسمت اعظمِ سیتوپلاسم را این رشتههای اکتینی پر کرده است.
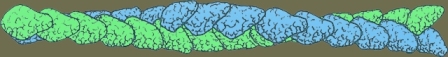
شکل: یک رشتۀ اکتین متشکل از هزاران پروتئین اکتین است که به صورت یک رشتۀ دو تاییِ در هم تابیده، شکل گرفتهاند.
میوزین نیز در سلولهای عضلۀ صاف به صورت رشتههایی طولانی در امتداد رشتههای اکتینی قرار گرفتهاند. در بالا ذکر شد که میوزین از سه ناحیۀ سر، بازو و دم تشکیل شده است. در عضلۀ صاف در هر رشتۀ میوزینی، هزاران میوزین از ناحیۀ دم به هم میچسبند و به صورت یک رشتۀ ضخیمتر در میآیند که از کنارههای این رشته، سرها و بازوها بیرون زدهاند. به این ترتیب از هزاران میوزین، یک رشته به وجود میآید که سرتاسرِ آن را سرها و بازوهای آمادۀ چسبیدن به اکتین پوشانده است.

شکل: میوزینهایی که در اتقباض عضلات نقش ایفا میکنند، در واقع از دو میوزین که دمِ آنها در هم تابیده شدهاست تشکیل شده.
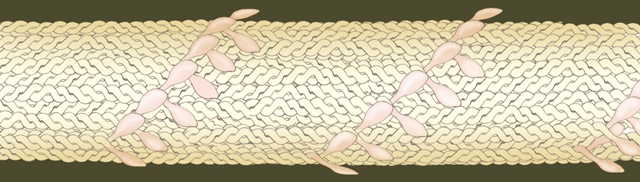
شکل: دمِ ملکولهای دوتاییِ میوزینها میتوانند در کنار هم قرار بگیرند و به موازاتِ یکدیگر، به هم بچسبند. به این ترتیب میتوانند رشتهای ضخیم را به وجود آورند که محدودیتی از لحاظ طول ندارد.
در میکروسکوپ الکترونی، قطر رشتههای میوزینی به وضوح از قطر رشتههای آکتینی ضخیمتر دیده میشود، به همین دلیل به رشتههای میوزینی، رشتۀ ضخیم (thick filament) و به رشتههای ساخته شده از آکتین رشتۀ نازک (thin filament) میگویند. میوفیلامان یا رشتۀ عضلانی نامی است که به هر دو این رشتهها داده شده است.
تا اینجا دانستیم که رشتههای نازک به عنوانِ ریل، و رشتههای میوزین که قدرتِ چسبیدن به اکتین و کشیدنِ آنها را دارد، در مجاورت هم همل سلولِ عضلانی صاف را پر کرده است. بنا براین تنها چیزی که لازم است تا نیروی خاصل از کششِ میوزینها به کلِّ سلول منتقل شود، این است که این رشتهها به محلهای محکم و ثابتی متصل شوند. این محلها نیز در سلول عضلانیِ صاف تعبیه شدهاند. در همسایگیِ غشاء و همچنین در میانۀ سلول، ساختارهایی ساخته شده از میوزین، اکتین و دیگر پروتئینها وجود دارند که به منزلۀ نقطۀ اتکاء برای رشتههای، ضخیم و نازک عمل میکنند. این ساختارهای محکم را “جسمِ سخت” (dense body) نامیدهاند. نیرویی که در اثر حرکتِ بازوها و چسبیدنِ “سر”های متعددِ میوزین به اکتین، به وجود میآید، به واسطۀ ثبات همین اجسامِ سخت، متمرکز میشود و به غشاء سلول عضلۀ صاف منتقل میشود و نهایتاً منجر به “انقباض” سلول میشود. انقباض همزمانِ صدها سلولِ عضلۀ صاف که در مجاورت هم هستند و به هم سخت چسبیدهاند نیز به نوبۀ خود منجر به انقباض یا کوتاه شدنِ رشتۀ عضلانی صاف میشود.
چگونگیِ اتصال رشتههای نازک و ضخیم به جسمهای سخت و ترکیب دقیقِ آنها و نیز کیفیت چسبیدنِ آنها به غشاء و سازکارِ انتقال نیرو توسط آنها به طور دقیق، شناخته شده نیست و هنوز موضوع تحقیق دانشمندان است [۴] – (Tajima-2008).
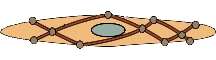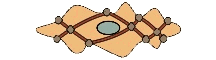
پویانما: یک سلول عضلۀ صاف؛ رشتههای آکتین و میوزین همۀ سلول را پر کردهاند و از این سو به آن سوی سلول کشیده شدهاند. همۀ این رشتهها به اجسام سخت در میان سلول و نیز در مجاورتِ غشاء سلول چسبیدهاند. اجسام سخت نیروی حاصل از تعاملِ میوزن و اکتین را متمرکز میکند و به غشاء سلول منتقل میکند و سلول منقبض میشود.
آرایش رشتههای اکتین و میوزین در عضلۀ مخطط منظمتر است. همۀ رشتهها در یک راستا و در کنار هم و به طور موازی سر تا سرِ تار عضلانی را آکندهاند. و به جای اجسامِ سخت که تکیه گاههای رشتهها برای اِعمال و انتقال نیرو هستند؛ در عضلۀ مخطط، مناطقی، با فواصل مساوی و کاملاً منظم هستند که رشتههای ضخیم و رشتههای نازک به آنها اتکا کردهاند. در مقطع عرضی، دیده میشود که رشتههای ضخیم، به فواصل مساوی از هم و با نظم کامل هندسی در کنار هم قرار گرفتهاند. در فاصلۀ بین رشتههای ضخیم نیز رشتههای نازک قراردارند، به طوری که هر رشتۀ ضخیم، به وسیلۀ ۶ رشتۀ نازک احاطه شده است. منظم بودن، همراستا بودن و دیگر نظامهایی که در تار عضلانی مخطط وجود دارد موجب میشود، انقباض در این عضله؛ ظریف و سریع و قابل کنترلتر باشد.

شکل: مقطع عرضی تار عضلانی و انتظامِ میوفیبریلها – این عکس به لطف و اجازۀ استاد دانشگاه واینونا آقای “بِرگ” از سایت آموزشی ایشان با تغییرات در نوشتهها مورد استفاده قرار گرفت. آیا میتوانید با استفاده از این عکس نسبت تعداد رشتههای نازک به تعداد رشتههای ضخیم را محاسبه کنید؟ اگر قطر یک میوفیبریل ۱ میکرومتر باشد تعداد تارهای ضخیم و تعداد تارهای نازک در آن چقدر است؟
{kind=link}
به این ترتیب سرهای میوزین که از پیرامونِ استوانۀ رشتۀ ضخیم بیرون زدهاند، با فاصلۀ کمی از اکتین قرار گرفتهاند و همواره ملکولِ اکتینی در دسترس دارند که در هنگام فرمانِ عصبی و آزاد شدنِ کلسیم به محیط، به آن بچسبند و چرخۀ انقباض را در سطح ملکولی آغاز کنند. در عضلۀ اسکلتی هر چند صد میوفیلامان، در یک گروه قرار گرفتهاند که توسط رتیکولوم سارکوپلاستیک احاطه شدهاند و تشکیل یک میوفیبریل را میدهند. هر میوفیبریل متشکل از حدود هزارو چند صد میوفیلامان است که با نظم فوق در کنار هم به طور فشرده قرار گرفتهاند. میوفیبریل سرتاسرِ تار عضلانی را طی میکند.
یک بار دوره کردنِ زیر مجموعههای یک عضله بی فایده نیست.
هر عضله از فاسیکولهای متعددی تشکیل شده. فاسیکولهای از سلولهای عضلانی (تار عضلانی) تشکیل شده. هر تار، متشکل از هزاران میوفیبریل است. میوفیبریل استوانۀ بسیار درازی است متشکل از صدها میوفیلامان. میوفیلامانها همان رشتههای ضخیم (=میوزین) و رشتههای نازک (=اکتین) هستند که به تناوب و با نظمی کامل، سارکومر را تشکیل میدهند.
هر میوفیلامان، توسط شبکۀ سارکوپلامی احاطه شده است.
هر تار عضلانی علاوه بر “صفحۀ پایه” توسط یک پردۀ نازک بافت پیوندی نیز احاطه شده است که به آن اندومیزیوم میگویند. بافت پیوندیِ دور یک فاسیکول عضلانی را پریمیزیوم و بافت پیوندی دور یک عضله را اپیمیزیوم مینامند. همۀ این مجموعۀ بافت پیوندی، به هم متصل هستند و تجمیع نیروهای ایجاد شده توسطِ سارکومرها، بر عهدۀ همین مجموعۀ بافت پیوندی است که در نهایت به تاندونها منتقل میشود.
معمولاً عضلات را به سه نوع صاف، اسکلتی و قلبی تقسیم بندی میکنند. دربرخی منابع، عضلۀ اسکلتی را مخطط مینامند. منشاءِ این نامگذاری، منظرهای است که زیر میکروسکوپ از این نوع عضله به چشم میآید. اما این منظره در عضلۀ قلبی نیز دیده میشود در حالی که سازمان سلولی عضلۀ قلبی با عضلاتی که در بدن به استخوانها متصل هستند آنچنان متفاوت است که از نامگذاری متمایز این دو گزیری نیست. بنا بر این بافت شناسان ترجیح دادهاند که از سه نوع عضله سخن بگویند. به این ترتیب یا باید نام عضلۀ صاف را عوض کنیم، یا منطقاً بپذیریم که دو نوع عضله داریم، صاف و مخطط و عضلۀ اسکلتی و عضلۀ قلبی را دو زیر شاخۀ عضلۀ مخطط بدانیم.
این عضله مشهورترین نوع عضله است. عضلات اسکلتی، مطابقِ نامش، متصل به اسکلت استخوانی بدن است. حرکاتِ آن ارادی و تحت فرمان دستگاه عصبی است. حرکتِ عضلۀ اسکلتی سریع و دقیق است. تارهای عضلانی این نوع عضله از هم از جهت الکتریکی از هم جدا هستند و به همین دلیل امکانِ این که هر یک از این اجزاء را به طور جداگانه تحریک کرد وجود دارد. در حالی که مثلاً در عضلۀ قلبی به این دلیل که سلولهای عضلانیِ آن به هم راه دارند و از هم عایق نشدهاند تحریک یک قسمت از این عضله، ناگزیر به همۀ آن سرایت میکند.
برخلاف بسیاری از بافتها، عضلۀ اسکلتی از سلولهای مجزا ساخته نشده است. بلکه از تارهای بسیار بزرگ چند سلولی ساخته شده است. این سولها نتیجۀ جوش خوردن تعداد زیادی سلول جنینی هستندئ که میوبلاست خوانده میشوند. در دیگر بافتها تکثیر سلولی موجب رشد در بافت میشود. در بافت عضلانی رشد به واسطۀ تکثیر هسته های موجود در تارها انجام میشود.
اگر چه تار عضلانی از جوش خوردن چندین سلول جنینی به وجود آمده و دارای تعداد بسیار زیادی هسته است اما معمولاً آن را سلول میخوانند؛ هرچند به واقع یک سلول نیست. قطر سلول عضلانی تقریباً در سراسرِ طولِ آن یکسان است و یک تار عضلانی معمولاً سرتاسرِ عضلهای را که در آن قرار دارد را طی میکند. به این ترتیب تار عضلانی یک استوانۀ بسیار طولانی تا حد چندین سانتیمتر و قطر حدود ۵۰ میکرومتر است.
سلولهای عضلۀ قلبی و عضلۀ صاف، در مقابل، متشکل از واحدهای سلولیی هستند که هر کدام، یک هسته دارند.
اصطلاحات بافت عضلانی
اجزاء سلولیِ بافت عضلانی را به نامهایی مخصوص به همین بافت میخوانند.
سیتوپلاسمِ تار عضلانی که انباشته از تارهای عضلانی است و جای زیادی برای دیگر اندامکهای سلولی باقی نگذاشته است راسارکوپلاسم؛ شبکۀ آندوپلاسمی را شبکۀِ سارکوپلاسمی و غشاء سلولی را سارکولم میخوانند. همچنین ممکن هنوز در برخی کتابها به میتوکندریِ تار عضلانی، سارکوزوم بگویند.
آنچه موجب شدهاست تا بافت شناسان همچنان از این نامهای اختصاصی برای اندامکهای سلول عضلانی استفاده کنند این است که این اندامکها در این نوع سلول به شدت تخصصی شدهاند و شکل و عملشان واقعاً با شکل و عمل نظائرشان در دیگر سلولها متفاوت است.
سلول عضلانی از معدود سلولهایی است که غشاء آن تحریک پذیر است. سلول عضلانی در این خاصیت فقط با سلول عصبی مشترک است. تغییر بار الکتریکی در قسمتی از این غشاء به سرتاسرِ ان سرایت میکند و از آنجا به شبکۀ سارکوپلاسمیک میرسد و در شبکۀ سارکوپلاسمیک موجب تخلیۀ ناگهانی کلسیم به داخل سیتوپلاسمِ تار عضلانی میگردد. تخلیۀ کلسیم اولین گام برای شروع عملیاتِ مشترکِ میوزن و آکتین است.
هستههای یک تار عضلانی، در وسطِ آن قرار ندارند بلکه به جدارِ تارها رانده شدهاند و تقریباً چسبیده به سارکولم قرار گرفتهاند. در مقطع عرضی در هر تار عضلانی یکی دوتا هسته ممکن است دیده شود اما در مقطع طولی تعداد این هستهها بسیار زیادتر خواهند بود.
در طول هر میوفیبریل، به طور منظم، و در فواصلِ مشخص، ساختارهایی تکرار میشود که به وسیلۀ میکروسکوپ نوری به صورتِ نوارهای روشنی که عمود بر تار عضلانی قرار گرفتهاند دیده میشوند. نام مخطط به خاطر وجود همین “خطوطِ” موازی به این نوع از عضلات داده شده است. این نوارها تمام طول میوفیبریل را پر کردهاند و مانند باسمهای در طول میوفیبریل تکرار میشوند. هر یک از این باسمهها، حدود ۲٫۵ میکرون طول دارد و سارکومر (واحد عضلانی) خوانده میشود. سارکومر به واقع کوچکترین ساختاری است که کارکردِ انقباض را بر عهده دارد. هنگام انقباض، طول سارکومر به ۱٫۵ میکرون کاهش مییابد و هنگام استراحت، به ۳ میکرون افزایش مییابد.
به وسیلۀ میکروسکوپ الکترونی، جزئیات بیشتری از این ساختار قابل مشاهده خواهد بود و ساختارهای زیر را میتوان از هم تشخیص داد:
خط میوزین (M-line) محلی است که رشتههای ضخیم از پشت به هم چسبیدهاند و از این نقطه این رشتهها به دو طرف امتداد یافتهاند.
خط اکتین (Z-line) محلی است که رشتههای اکتین در آنجا به هم متصل شدهاند و سرِ آزادِ آنها به سمتِ خط میوزین جهت یافتهاند.
نوار همسان (I-bamd) قسمتی است که فقط رشتههای نازک در آن ناحیه قرار دارد
نوار ناهمسان (A-band) قسمتی است که رشتۀ ضخیم متشکل از میوزین آن را اشغال کرده اما در قسمتی از آن، رشتههای نازک هم حضور دارند. در ناحیۀ نوار ناهمسان، منطقهای که هر دو رشته وجود دارند، تیره تر دیده میشود. قسمتی از این نوار که رشتههای ضخیم به تنهایی وجود دارند، روشنتر از بقیۀ ناحیۀ نوار ناهمسان است و به همین دلیل، نوار روشن (H-band) نامیده شده است؛ اگر چه روشنی این ناحیه از نوار همسان بیشتر نیست.
کشف این نواحی اولین بار به وسیلۀ دانشمندان آلمانی صورت گرفته و نامگذاری نواحیِ مختلف در عضلۀ مخطط، به احترام آنان در متون غیر آلمانی نیز به همان نامهای آلمانی خوانده میشود. نویسندگان ایرانی نیز در کتابهای خود نام جدیدی بر این نواحی نگذاشتهاند.
تیتین از خط میوزین تا خط اکتین امتداد یافته قسمتی از تیتین حالتی کشسانی دارد و انتهای این قسمت به خط اکتین متصل است. تیتین به این ترتیب در حفظِ انتظامِ رشتههای عضلانی ایفاء نقش میکند و همچنین به همراه بافت همبند، موجب بازگشت عضله به طول معمول خود در حالت عادی میشود.
مولکولهای دیگری نیز خطهای آکتینِ میوفیبریلهای مجاور را به هم متصل میکند. میوفیبریلهایی که در اطراف تار عضلانی قرار دارند نیز به وسیلۀ پروتئینهای دیگر به سارکولم یا غشاء سلول متصل هستند. به این ترتیب ثباتِ سازمانِ سارکومرها و انتقال مستقیم نیروی تولید شده در آنها به دو طرفِ تار عضلانی تضمین میشود.
هر تار عضلانی، توسط یک آکسونِ حرکتی تعصیب میشود. این آکسون ممکن است تارهای دیگر عضلانی را نیز عصب دهد. همۀ تارهای عضلانیی که توسط یک آکسون تعصیب میشوند را یک واحد حرکتی میخوانند. تعداد تارهای عضلانی یک واحد حرکتی در اندامی که حرکات ظریف و دقیق دارد (مثل عضلات چشم)، کم و تعداد تارهای واحدهای حرکتی که حرکات زمخت دارند (مثل حرکات ران و پشت) زیادتر است. پایانۀ آکسون به میانِ تار عضلانی میرسد و در آنجا پهن میشود و به صورت یک صفحه تغییر شکل میدهد. به این قسمت، صفحۀ انتهایی حرکتی یا اتصال عصب-عضله میگویند. صفحۀ انتهایی، معادلِ سیناپسِ بین نورونی است و در همین جا است که عصب نوروترانسمیتر یعنی استیل کولین را در فاصلۀ بین غشاء عصب و غشاء سلولیِ تار عضلانی ترشح میکند. گیرندههای موجود در غشاء تار عضلانی در صفحۀ انتهایی پس از اتصال با استیل کولین، موجب ایجاد پتانسیل عمل در غشاء تار عضلانی میشوند که با سرعت، در سراسرِ غشاء تار عصبی میپراکند. وجود آوندکهای عرضی موجب میشود که پتانسیل عمل همزمان تا عمق سلول نیز پیشرفت کند.
با توجه به آنچه شرح دادیم، میوفیبریل، یک سازمانِ کامل و آماده برای انقباض است. میوزینها با تراکم بالا در کنار ملکولهای اکتین قرار گرفتهاند و تعداد بسیار زیادی “سَر” در مقابل مولکولهای اکتین قرار گرفته است. اما قسمتهایی از اکتین که تمایل به چسبیدنِ به “سر”های اکتین را دارند توسط ملکولهای دیگری (تروپومیوزین) پوشانده شدهاند. در واقع تنها مانع برای این که “سر” میوزینها به اکتین بچسبد و چرخۀ انقباض آغاز شود همین تروپومیوزین است. آن چه میتواند موجب شود که تروپومیوزین جابه جا شود و محلِ چسبنده به میوزین را برهنه کند، یون کلسیم و اتصال این یون به تروپونین است که این تروپونین به نوبۀ خود به تروپومیوزین چسبیده است.
یون کلسیم به مقدار فراوان در شبکۀ سارکوپلاسمی ذخیره شده است. برای آن که در لحظۀ فرمانِ انقباض از جانب آکسون، کلسیم به مقدار کافی و به فوریت در اختیار همۀ تروپونینها قرار گیرد، این شبکه، سرتاسر سطحِ میوفیبریل را پوشانیده و احاطه کرده است. شبکۀ سارکوپلاسمیک در سلول عضلانی، مانند کیسهای همۀ فاصلههای میان میوفیبریلها را پر کرده و همۀ سطوحِ میوفیبریلها را احاطه کرده و فراگرفته و آماده است تا به محض رسیدنِ فرمان، ذخیرۀ غنی کلسیمی خود را بیرون بریزد و در اختیار تروپونینِ روی اکتین قرار دهد. گستردگی این شبکه در سراسر سلول عضلانی و در مجاورت همۀ میوفیبریلها موجب میشود که پس از صادر شدن دستور، همۀ واحدهای انقباضی (سارکومرها) با هم و در یک زمان، کلسیم لازم را در دسترس داشته باشند. شبکۀ اندوپلاسمی، با چنین شکل و انتظامی در میان همۀ سلولهای بدن مختصّ سلول عضلانی است. شاید به همین دلیل باشد که بافت شناسان ترجیح میدهند نام متمایزِ شبکۀ سارکوپلاسمی را به این اندامک در سلول عضلانی بدهند.
به این ترتیب همه چیز آماده است و تنها چیزی که برای آغاز انقباض لازم است این است که غشاء این شبکۀ سارکوپلاسمی تحریک شود. اما در اینجا آنچه مهم است این است که همۀ سطحِ این شبکه، یکجا و در یک زمان با هم تحریک شود. تنها در این صورت است که این شبکه میتواند کلسیم ذخیره شده در خود را در یک زمان، در اختیار همۀ میوفیبریلهای سراسرِ طولِ سلولِ عصبی قرار دهد. آنچه این تحریک سرتاسریِ همزمان را تضمین میکند، شکل خاصی است که غشاء سلولیِ این سلول دارد.
همانطور که گفتیم، غشاء سلول عضلانی یک غشاء تحریک پذیر است. سارکولِم این قابلیت را دارد که پالس الکتریکی به صورت پتانسیل عمل، در سطحِ آن منشر میشود. سارکولم در سطح خود فرورفتگیهای متعددی دارد که به صورتی کاملاً منظم قرار دارند. هر یک از این فرورفتگیها دهانۀ یک آوندک هستند و این آوندکهای نازک و طولانی تا اعماق سلول پیش رقتهاند و در امتداد خطِ اکتین، حولِ میوفیبریل را احاطه کردهاند. به نحوی که هر جا خطِ اکتینی هست، یک حلقه از جنس و در ادامۀ غشاء سلولی، هم هست. این غشاء اگر چه تا اعماق سلول پیش رفته اما این غشاء محل عبور مادهای از خارج سلول به داخل سلول نیست بلکه صرفاً موجب میشود که دِپلاریزاسیون ایجاد شده در قسمتِ سطحیِ سلول بتواند به سرعت به داخلِ غشاء منتشر شود. در دو طرف و در همسایگیِ هر یک از این آوندکها یا حلقههای غشائی، شبکۀ سارکوپلاسمی قرار دارد. به این ترتیب امکان تحریک یکباره و همزمان همۀ شبکۀ سارکوپلاسمی فراهم میشود.
شبکۀ سارکوپلاسمی در سرتاسرِ سطحِ میوفیبریل گسترده شده است اما در همسایگی آوندک متسع شده و به صورت دو آوندکِ دیگر البته از جنسِ شبکه سارکوپلاسمی شکل گرفته است که به آن دو نامِ کیسۀ شبکۀ سارکوپلاسمی دادهاند. شباهتِ این آوندکها و همجواریِ آنها موجب شده که به این مجموعه نامِ “سهقلو” بدهند. خصوصاً منظرۀ این ساختار در میکروسکوپ الکترونی، به صورتِ سه لولۀ چسبیده به هم نامِ سه قلو را شایستۀ این ساختار میکند.
در عضلات اسکلتی ساختاری تخصصی به نام دوک عضلانی وجود دارد که کشش عضله را دریافت میکند و “حس عمقی” نتیجۀ کارکرد آن است. دوک عضله، متشکل از یک دسته از سلولهای عضلانی کوتاه یا هستههای مرکزی است که درون یک پردۀ کلاژنیِ دوکی شکل قرار گرفته و از بقیۀ بافت عضلانی جدا شده است. کشیده شدنِ عضله منجر به کشیده شدنِ این دوکها میشود و کشیده شدنِ دوک، جرقۀ شروعِ بازتابِ کشش است.
دوک عضلانی تنها پایانهای نیست که متولی حس عمقی است. پایانههای مشابهی در تاندون و مفصل نیز وجود دارند که کارکرد مشابه دارند.
چنان که دیدیم، سازکارهای متعددی که در غشاءِ تار عضلانی و شبکه سارکوپلاسمی و میوفیبریلها تعبیه شده و معماری منظم و خاصی که اجزاء محتلف تار عضلانی دارد موجب میشود که انقباض در سراسر یک تار عضله، همزمان انجام شود و این همزمانی است که سرعت، دقت و هماهنگی را در حرکات عضلۀ اسکلتی تضمین میکند. روندی که در آن دپلاریزاسیونِ سارکولم، به انقباضِ میوفیبریلها منجر میشود را جفت شدنِ تحریک- انقباض میگویند. ما در بالا این روند را تشریح کردهایم و در اینجا یک بار دیگر کل این روند را دوره میکنیم؛
۱) ترشح استیل کولین از صفحۀ انتهایی، موجب دِپلاریزاسیونِ سارکولِم در این ناحیه و ایجاد پتانسیل عمل میشود. ۲) پتانسیل عمل، در سراسرِ غشاءِ سلول منتشر میشود. آوندکهای عرضی ادامۀ غشاء هستند، آوندکها در عمق سلول نفوذ کردهاند و دورِ میوفیبریلها حلقه زدهاند و در دو طرفِ هر آوندک، دو کیسۀ سارکوپلاسمی قرار گرفته است. ۳) دپلاریزاسیون به آوندکهای غشائی منتشر میشود. ۴) تبادلِ یونی میانِ غشاء آوندکها و غشاءِ کیسههای سارکوپلاسمی اتفاق میافتد و این منجر به تخلیۀ یکبارۀ کلسیم از شبکۀ سارکوپلاسمی در داخل سارکوپلاسم میشود. ۵) کلسیم به تروپونین میچسبد و موجب تغییر شکل فضاییِ آن میشود. مناطقی از رشتۀ اکتین که تمایل به چسبیدن به میوزین دارد برهنه میشود. ۶) سرهای میوزینها به اکتین میچسبد. ۷) بازوی میوزین به تحریک یونِ منیزیم و با مصرفِ آ.ت.پ. و تبدیلِ آن به آ.د.پ. خم میشود. و رشتۀ نازک (اکتین) به اندازۀ چند نانومتر به رشتۀ ضخیم (میوزین) و در واقع به خط میوزین نزدیک میشود. ۸) یک آ.د.پ. دیگر به میوزین میچسبد و موجب آزاد شدن و باز شدنِ مجددی بازوی میوزن میشود.
حضورِ کلسیم و آ.ت.پ. کافی در دسترس میوفیلامانها موجب تکرار این چرخه میشود.
آنچه در واقع اتفاق میافتند بسیار پیچیدهتر از آن است که در بالا گفته شد. پروتئینهای متعدد دیگری در این روند دخالت دارند که این روند را بسیار پیچیده و تخصصیتر میکنند. در برخی از مراحل این روند از جمله کیفیتِ انتقال و ترجمۀ پیام از سارکولم به کیسههای شبکۀ سارکوپلاسمی، موضوعِ فعلیِ تحقیقِ محققان است.
هر سلولِ عضله، توسطِ اندومیزیوم که پردهای نازک از تارهای کلاژن است احاطه شده که در واقع آن را میتوان عضوی از بافت پیوندیِ عضله دانست. درون اندومیزیوم، آکنده از مویرگ است که کارکردِ همیشگی خود در انتقال گازهای تنفسی و مواد غذایی را بر عهده دارد. هر تار عضلانی متکی به صفحۀ پایه است و به توسطِ این صفحه با اندومیزیوم متصل است. اندومیزیوم، نیروی حاصل از انقباض تارهای عضلانی را به دو انتهای تار عضلانی منتقل میکند. انتهای همۀ اندومیزیومهای یک عضله در انتهای عضله به هم متصل هستند و در انتهای عضله تنگ در هم تنیده و چسبیدهاند و تشکیل تاندون یا فاسیا را میدهد. این ساختار موجب میشود که نیروی انقباضیِ همۀ میوفیبریلها در دو انتهای عضله تجمیع شود و یکجا به استخوان منتقل گردد. ساختار و تعاملِ تارِ عضله، صفحۀ پایه و اندویزیوم و تارهای کلاژنیِ آن خصوصاٌ در دو انتهای تار عضله اهمیت مییابد. در این منطقه، صفحۀ پایه مضرس میشود و استطالههای آن به درونِ منطقۀ تاندون نفوذ میکند. همچنین، کلاژنها به سوی متقابل، نفوذ کردهاند. غشاء خودِ تار عضله نیز، ساختارهای اتصالیِ سختی با صفحۀ انتهایی برقرار میکنند. به این ترتیب انتقال نیرو از میوفیبریلها به تاندون و از آن طریق به استخوان تضمین میشود.
چنان که میبینیم، بافت همبند در عضله نیز کارکردِ مرسومِ خود را دارد و در اینجا علاوه بر این که جایگاه عبور عروق خونی است، متخصص در انتقال نیروی مکانیکی است.
در همسایگی تارهای عضلانی، سلولهای اقماری نیز دیده میشود که احتمالاً سلولهای بنیادینی هستند که قابلیت تبدیل به تار عضلانی را دارند. با استفاده از میکروسکوپ نوری و رنگامیزی هماتوکسیلین ائوزین تشخیص و تمایز هستۀ این سلولهای اقماری، سلولهای اندوتلیال، سلولهای فیبروبلاستِ اندومیزیوم و سلولهای عضلانی همیشه آسان نیست.
با نظامِ رشتههای اکتین و میوزین در عضلۀ صاف آشنا شدیم. در اینجا خصوصیات بافت شناختی آن را دوره میکنیم. این نوع عضله بر خلاف عضلۀ اسکلتی از سلولهای مجزا با هستههای یگانه تشکیل شده.
کارکرد عضلۀ صاف با آنِ عضلۀ اسکلتی متفاوت است. حرکت در عضلۀ اسکلتی فوراً ایجاد میشود و سریعاً نیز خاتمه مییابد. امکانِ انقباض طولانی مدت در عضلۀ اسکلتی وجود ندارد. در عضلۀ اسکلتی امکان انقباض طولانی هست. این خصیصه برای حفظ انقباض در عروق و دستگاه گوارش ضروری است. انقباض برخی از اسفنکترهای عضلانیِ صاف، میتواند تا یک روز ادامه یابد.
اگر عضلۀ اسکلتی بیش از حد کشیده شود، قدرتِ کافی نخواهد داشت، در واقع اگر یک عضلۀ اسکلتی را آنقدر بکشیم تا طولش به حدود دو برابر اندازۀ عادی خود باشد، هیچ سرِ میوزینی با هیچ رشتۀ اکتینی تماس نخواهد داشت و اساساً امکان اِعمالِ نیرو در آن وجود نخواهد داشت. انعطافی که در اندامهای مجهز به بافت صاف عضلانی وجود دارد موجب میشود که عضلۀ صاف در طولهای مختلف قابلیت انقباض خود را از دست ندهد. اگر عضلات دیوارۀ معده از نوع مخطط میبودند، هنگامی که معده از غذا پر میشد عضلاتِ دیوارۀ آن دیگر امکان عمل نمیداشتند.
عضلۀ صاف به جز سیستم عصبی تحت تأثیر دیگر عوامل از جمله عوامل موجود در محلِ استقرار عضله نیز منقبض میشود. این خصلت موجب میشود که اندامهای مجهز به عضلۀ صاف بتوانند، مستقل از دستگاه عصبی و نیز در پاسخ به تغییرات محلی عمل کنند. اگر در خواب باشید غذا همچنان در روده حرکت میکند، و فشار خون همچنان تنظیم میشود و اگر قسمتی از بدن به خون بیشتری نیاز داشته باشد، خون کافی به آن میرسد. اینها همه هنرِ عضلۀ صاف است. عضلۀ اسکلتی فقط از دستگاه عصبی دستور میگیرد و لا غیر. به هر حال حرکت عضلات صاف اگر چه تحت تأثیر دستگاه عصبی (خودکار) باشد؛ غیر ارادی است.
|
اگر حیوانی را تشریح کنید، میتوانید این خاصیت عضلۀ صاف را به وضوح در رودههای حیوان مشاهده کنید. هنگامی که حیوان عمیقاً بیهوش است، اگر قسمتی از رودۀ آن را به طور مکانیکی تحریک کنید مثلاً با دو سرِ انبرک یا هر وسیلۀ دیگری آن را فشار دهید، یک انقباض موضعی در محل ایجاد میشود که به صورت حرکت دودی ادامه مییابد. این تحریک با مواد شیمیایی مثل نیکوتین نیز ایجاد میشود. دیدن این انقباض و حرکتِ دودی مخصوصاً وقتی جالبتر است که حیوان مرده باشد و قلب، ریه و مغز حیوان کاملاً از کار افتاده باشد. اگر مدت زیادی از مرگ حیوان نگذشته باشد، همچنان میتوان اثر این تحریک را دید. |
لازمۀ دوام و طولانی بودنِ انقباض در عضلات صاف، مصرفِ کمتر انرژی در آن است. عضلۀ صاف بسیار کم مصرف تر از عضلۀ مخطط است.
تمرکز در اعمال نیروی مکانیکی و سرعت و دقت، در عضلۀ صاف، قربانیِ دوام و کم هزینگی انقباض شده است. در عضلۀ مخطط، کم هزینگی و دوام، قربانی سرعت و تمرکز و دقت شده است. به همین دلیل است که وجود عضلۀ مخطط بدون استخوان یا بافت سختی شبیه آن بسیار نادر است.
سلولهای عضلانی صاف به وسیلۀ اتصالِ روزنه به هم مربوطند به این ترتیب تحریک یک سلول به بقیۀ سلولهای مجاور سرایت میکند. در عضلۀ صاف ساختاری چون صفحۀ انتهایی وجود ندارد.
هستۀ سلول در عضلۀ صاف کشیده و دوکی شکل و به اصطلاح بافت شناسان شبیه سیگار برگ است. طول سلول حدود ۵میکرون است اما در برخی موارد تا ۱۰۰ میکرون و بیشتر هم میرسد (طول یک سلول عضلۀ صاف در رحم زن حامله به ۶۰۰ میکرون میرسد) اما به هر حال بسیار کوتاه تر از سلول عضلۀ اسکلتی است.
عضلۀ صاف را میتوان به دو گروه عضلات صاف چند واحدی و تک واحدی تقسیم کرد. در نوع دوم که به عضلۀ صاف احشایی نیز خوانده میشود، همۀ عضلات مجاور به صورت یک واحد عمل میکنند و قسمتهای مجاور استقلال عمل از هم ندارند.
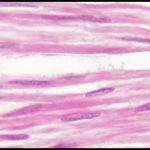 زیر میکروسکوپ مقطع طولی عضلۀ صاف هستههای کشیده را نمایان میسازند. حدود سلولها به سختی قابل تشخیصاند. خطوطی که در عضلۀ مخطط دیده میشوند در اینجا وجود ندارند. گاهی با بافت همبند متراکم منظم اشتباه میشوند. اما هستۀ فیبروبلاست، نازکتر و کشیدهتر است، بافت همبند، نامنظمتر است. هسته در بافت عضلات صاف، در کنار و درونِ تارها (میوفیبریلها) قرار دارند در حالی که هستۀ فیبروبلاست خارج و جدا از تارها (کلاژن) قرار دارند. هستهها در بافت همبند، در جهات متفاوت و با فواصلِ متفاوت قرار گرفتهاند. هستۀ سلول عضلۀ صاف در حال انقباض فنری شکل است.
زیر میکروسکوپ مقطع طولی عضلۀ صاف هستههای کشیده را نمایان میسازند. حدود سلولها به سختی قابل تشخیصاند. خطوطی که در عضلۀ مخطط دیده میشوند در اینجا وجود ندارند. گاهی با بافت همبند متراکم منظم اشتباه میشوند. اما هستۀ فیبروبلاست، نازکتر و کشیدهتر است، بافت همبند، نامنظمتر است. هسته در بافت عضلات صاف، در کنار و درونِ تارها (میوفیبریلها) قرار دارند در حالی که هستۀ فیبروبلاست خارج و جدا از تارها (کلاژن) قرار دارند. هستهها در بافت همبند، در جهات متفاوت و با فواصلِ متفاوت قرار گرفتهاند. هستۀ سلول عضلۀ صاف در حال انقباض فنری شکل است.
با این وجود امکان اشتباه بافت همبند با عضلۀ صاف خصوصاً هنگامی که اُریب بریده شده باشد هست.
اگر در یک نمونۀ میکروسکوپی در تشخیص قسمتی از آن به عنوانِ بافت همبند یا عضلۀ صاف دچار اشکال شویم، یافتنِ یک شریان در همان نمونه راهگشا خواهد بود. در دیوارۀ شرائین هر دو این بافتها وجود دارد. در میانِ دیوارۀ شریان (مدیا) عضلۀ صاف و در خارجِ آن (ادوانتیس). مقایسۀ بافت مشکوک، با این دو قسمت از شریان تشخیص شما را به واقعیت نزدیکتر خواهد کرد.
قلب توسط اعصاب خودکار تحریک میشود و یک عضلۀ مخططِ غیرارادی است. بنابراین از یک نظر شبیه عضلۀ اسکلتی و از نظری شبیه عضلۀ صاف است. این عضله از سلولهای مجزا با هستۀ یگانه ساخته شده است. سلولهای عضلۀ قلبی از دو انتها، به سلولهای مجاور از طریق قرصۀ بینابینی متصل هستند. قرصۀ بینابینی، حاوی اتصال داربستی چسبنده و اتصال روزن است. ترکیب این دو نوع ساختار به همراه هم در بدن نادر است و در عضلۀ قلبی، دو کارکرد را در پی دارد. تجمیع نیروهای مکانیکی و اتحادِ الکتریکی. اتصال داربست موجب انتقال نیروی مکانیکی ایجاد شده در یک سلول به سلول مجاور میشود و اتصال روزن با ایجاد امکان تبادل یونها میان سلولهای مجاور، اتصال الکتریکی آنها را تأمین میکند.
سرعت انتقال پیام الکتریکی، به توسطِ انتقال یونها از طریقِ قرصههای بینابینی، سرعتِ متوسطی دارد و موجب میشود که انقباضِ آغاز شده از بالای قلب، به تدریج و با فاصلۀ زمانی به پایین سرایت کند. برای این که راندمان کار قلب در اخراجِ خون از بطن بالا رود طنابهای موسوم به تارهای پورکنژ تعبیه شدهاند که متخصص انتقال پیام الکتریکی هستند و حکم میانبر برای پیام الکتریکی به قسمتهای مختلف عضلۀ بطنی را دارند. تحریک الکتریکی عضلۀ قلب از گره سینوسی، دهلیزی آغاز میشود. به این ترتیب انقباض دهلیز، از بالا آغاز میشود و به پایین سرایت میکند و همزمان، گره دهلیزی بطنی تحریک میشود. اما جسمِ فیبری مانعِ عبور جریان الکتریکی از طریق دهلیز به بطن میشود. موج الکتریکی به این ترتیب از طریق طناب پورکنژ به همۀ بطن راه مییابد. سرعتِ موج در این طناب ۴ تا ۵ برابر آن در خود عضله است.
سلولهای اقماری در قلب وجود ندارند و به این جهت تکثیر د رآنها اتفاق نمیافتد.
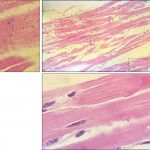 زیر میکروسکوپ نوری؛ هستۀ سلولهای عضلانی، بیضوی، نسبتاً کمرنگ و در مرکز سلول دیده میشوند. تخطیط در آنها مثل عضلۀ اسکلتی دیده میشود. اما نظم خطوط عرضی در این عضله به جهت وجود قطرات چربی و دانههای گلیکوژن کمتر است. در اطراف هستۀ سلول سیتوپلاسم کمرنگتر دیده میشود. این منطقه خالی از میوفیبریل است.
زیر میکروسکوپ نوری؛ هستۀ سلولهای عضلانی، بیضوی، نسبتاً کمرنگ و در مرکز سلول دیده میشوند. تخطیط در آنها مثل عضلۀ اسکلتی دیده میشود. اما نظم خطوط عرضی در این عضله به جهت وجود قطرات چربی و دانههای گلیکوژن کمتر است. در اطراف هستۀ سلول سیتوپلاسم کمرنگتر دیده میشود. این منطقه خالی از میوفیبریل است.
قرصههای بینابینی گاهی کمرنگتر و گاهی پررنگتر از سیتوپلایم و به صورت پلهای دیده میشوند.
تارهای پورکنژ کمرنگتر از سیتوپلاسمِ سلولهای عضلانی دیده میشوند و قطورترند.
پانوشت:
[۱] http://en.wikipedia.org/wiki/Fastest_animals اگر سرعت را نه با “متر بر واحد زمان” بلکه با “طول بدن” بر واحد زمان بسنجیم، شاهین حیوان کم سرعتی خواهد بود و احتمالاً باکتریهای فلاژل دار صدر نشینِ این مسابقه خواهند بود.
[۲] http://www.nature.com/scitable/knowledge/library/animal-migration-13259533
[۳] D. P. Syamaladevi, J. A. Spudich, and R. Sowdhamini, “Structural and functional insights on the Myosin superfamily,” Bioinformatics and Biology Insights, vol. 6, pp. 11–۲۱, ۲۰۱۲٫
دسترس در http://www.ncbi.nlm.nih.gov/pubmed/22399849 در دیماه ۱۳۹۶
[۴] Tajima Y, Takahashi W, Ito A (2008) Small-angle X-ray diffraction studies of a molluscan smooth muscle in the catch state. J Muscle Res Cell Motil 29:57–۶۸٫ doi:10.1007/s10974-008-9142-0
دسترس در http://link.springer.com/article/10.1007%2Fs10974-008-9142-0?LI=true در دیماه ۱۳۹۶
سلام و خسته نباشید
اما مانع اتصال اکتین و میوزین ، تروپومیوزین است نه تروپونین!!!!!
عزیزِ گرامی سیامک
از تذکرت بسیار سپاسگزارم. پس از ملاحظۀ تذکر شما اصلاحات لازم را بی درنگ اِعمال کردیم. این صفحه تا وقتی خوانده شود مرهون لطف و دقت و توجه شما خواهد بود.
سلام و خسته نباشید
اما مانع اتصال اکتین و میوزین ، تروپومیوزین است نه تروپونین!!!!!
خسته نباشید ، الان در انقباض ماهیچه طول ناحیه H سارکومر به حداکثر میرسه؟
خسته نباشید ، الان در انقباض ماهیچه طول ناحیه H سارکومر به حداکثر میرسه؟
ممنون از وبلاگ عالیتون
ببخشید دو یارکومر کنار هم دقیقا چند خط سفید دارند؟
ممنون از سایت خود و مفیدتون
کدام ماهیچه های اسکلتی به صورت جفت عمل نمیکنند؟؟
اسکلت خارجی چه کمکی به حرکت میکند؟
منظور شمااز جفت شدن اگر “کوپلینگ” (coupling)باشد، هیچ عضله ای نیست که بدون این مکانیسم منقبض شود.
اگر منظورتان “آنتاگونیسم” (antagonism) است. باید گفت که باید تک تک عضلات را بررسی کرد تا ببینیم آیا عضله ای وجود دارد که آنتاگونیست نداشته باشد. شاید بشود عضلۀ قلب را عضله ای دانست که آنتاگونیست ندارد. هنگامی که عضلۀ قلبیِ بطن منقبض میشود، هیچ عضلۀ دیگری نیست که آن را به حالت قبلی خود باز گرداند بلکه این فشارِ خونِ ورودی به قلب است که بطن را به حال اولیه خود بر میگرداند.
ویژگی خط mدر بافت اسکلتی چیست
سلام
ای کاش عکس بافت های ماهیچه ی قلبی و صاف رو هم میذاشتین
:)
حامد عزیز
از تذکر شما سپاسگزاریم
طی دو هفتۀ آینده به خواست خداوند عکسهایی از این بافتها خواهیم گذاشت.
سلام میشه بگین کدوم ماهیچه انقباض کند و طولانی داره؟
صاف ^_^